発達生理学研究系
概要
発達生理学研究系は,平成15年度より「心と身体の発達における生理学的機構をあきらかにすること」を目的として,専任3部門,客員1部門の研究系として発足した。専任1部門はそれまで統合生理研究施設高次脳機能研究プロジェクト(伊佐正教授)が配置換えで認知行動発達機構研究部門として発足し,眼球や手の運動を制御する神経回路の発達と損傷後の機能代償機構について研究を展開している。また生体恒常機能発達機構研究部門の教授には鍋倉淳一教授が選出され,平成15年より中枢神経系のシナプスの発達および脳損傷後の神経回路のリモデリングのメカニズムを電気生理学的研究手法や多光子顕微鏡などを用いて解析している。また生殖内分泌機能発達研究部門には箕越靖彦教授が選出され,平成15年に着任し,肥満の原因となる視床下部の代謝中枢に関する研究を行っている。また客員研究部門については,椛秀人教授(高知大学医学部)が5年間環境適応機能発達研究部門の客員教授として勤めていただいたが,平成20年度末で任期満了となり,現在後任人事の選考中である。
認知行動発達機構研究部門では脳による運動制御,特に眼球のサッケード運動系及び手指の物体把持(grasping)運動について大規模及び局所神経回路の構造と機能の解析,さらには損傷時の機能代償機構を研究している。
1.特に眼球のサッケード運動系については
(1)サッケードの制御にとって重要な中枢である中脳上丘の局所神経回路の構造と機能をマウスの脳スライス標本においてパッチクランプ法などを用いて解析する。
(2)麻酔下動物(ラット,サル)における上丘,大脳皮質をめぐる大規模神経回路及び上丘局所神経回路のシナプス機構を解析する。
(3)遺伝子改変マウスにおいてサッケード運動を解析する事を通じてサッケード制御回路の分子機構の明らかにする。
(4)覚醒サルにおける電気生理学,薬物微量注入法を用いてサッケード制御系の動的特性を解析する。
(5)大脳皮質一次視覚野(V1)を損傷したサルを,いわゆる「盲視」の動物モデルとして用い,V1損傷後も障害視野へのサッケード運動を可能とする神経機構の解析,および盲視における「見え(視覚的意識)」とその神経機構を明らかにする。
2.手指の運動については,
最近我々は霊長類において大脳皮質一次運動野から手指筋運動ニューロンに至る経路には直接結合のほかに頚髄の介在ニューロンを介する間接的な経路も存在することを明らかにした。そして中部頚髄レベルで直接結合を遮断し,間接経路のみを残したサルにおいても1‐3ヶ月の回復期を経て手指の精密把持運動が回復になることを見出した。このモデルを用いて,機能代償の神経機構をPETによる脳活動イメージング,電気生理学,神経解剖学,遺伝子発現解析など様々な手法を解析している。また手や指の運動の正確な制御を可能にする脊髄神経機構を明らかにするため,覚醒行動中のサルの脊髄からの神経活動記録などを行って解析している。
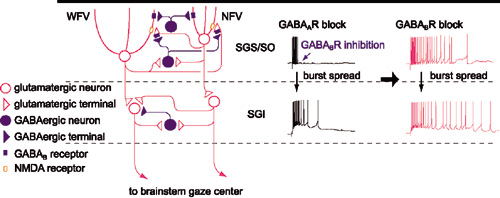 |
図1 上丘でのGABAB受容体を介したバースト発火の制御機構の模式図。GABAB受容体は上丘浅層のnarrow field vertical cell (NFV)とwide field vertical cell (WFV) のシナプス後膜およびグルタミン酸作動性神経終末に発現している。浅層のGABA作動性ニューロンのバースト発火によって遊離されたGABAが,これらの受容体に作用するとNFVでは過分極が,WFVではシャンティング抑制が誘発され,また,シナプス終末からのグルタミン酸遊離が抑制される。
|

|
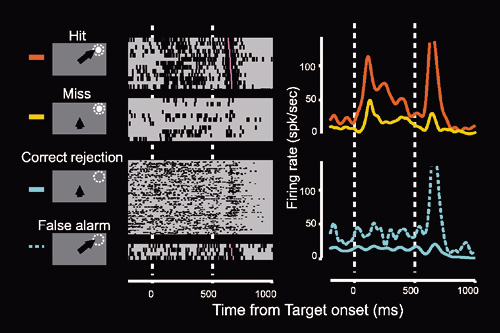 |
図2 片側一次視覚野除去後のサルから計測された,視覚刺激検出課題遂行中の損傷側上丘の神経活動。損傷側上丘・中間層の神経細胞ではサルの行動と相関した視覚応答の変化が見られる。ターゲットの検出・サッケードによる定位に成功した試行(Hit trial)では明確な視覚応答が観察されるのに対し,サルがターゲットを検出できなかった試行(Miss trial)では,視覚刺激はまったく同じであるにもかかわらず視覚応答は減衰している。
|
|
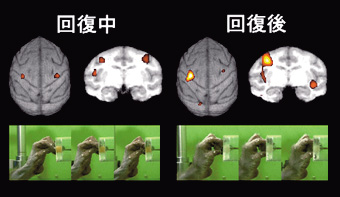 |
図3 頸髄レベルでの皮質脊髄路損傷後の機能回復初期(1ヶ月;左)と回復安定期(3ヶ月;右)の手指の運動(写真)と運動遂行中の大脳皮質の活動増加部位(図左が手の反対側)。初期は両側の一次運動野,安定期には反対側一次運動野と両側の運動前野の活動が増加する。
|
職 員
 |
教 授 伊 佐 正 ISA,Tadashi
東京大学医学部卒,同医学系研究科修了,医学博士。スウエーデン王国イェテボリ大学客員研究員,東京大学医学部助手,群馬大学医学部助教授を経て平成8年1月から現職。
専攻:神経生理学。
|
 |
助 教 関 和 彦 SEKI,Kazuhiko
新潟大学教育学部卒,筑波大学医学系大学院博士課程修了,医学博士。国際武道大講師,米国Washington大学客員研究員を経て平成13年4月から現職。
専攻:神経生理学,運動生理学。
|
 |
助 教 吉 田 正 俊 YOSHIDA, Masatoshi
東京大学薬学部卒,東京大学大学院薬学系専攻博士課程中退,医学博士。科学技術振興事業団心表象プロジェクト研究員,科学研究費(特別推進)研究支援員を経て平成15年6月から現職。
専攻:認知神経科学,神経生理学。
|
 |
助 教 金 田 勝 幸 KANEDA, Katsuyuki
京都大学薬学部卒,同薬学研究科博士課程修了,薬学博士。東京都神経科学総合研究所客員研究員,テネシー大学ポスドクを経て平成17年9月から現職。
専攻:神経生理学,神経薬理学。
|
 |
研究員 渡 辺 秀 典 WATANABE, Hidenori
東京理科大学理学部第二物理学科卒,東京大学工学系研究科修士課程,同大学院博士課程修了,工学博士。玉川大学学術研究所研究員,日本学術振興会特別研究員,玉川大学学術研究所OCE助手,同大学脳科学研究所特別研究員を経て平成20年から現職。
専攻:神経科学,生体情報システム工学。
|
 |
研究員 坂 谷 智 也 SAKATANI, Tomoya
東京大学理学部卒,同大学院修士課程修了,総合研究大学院大学博士課程修了,博士(理学)。生理学研究所研究員,オックスフォード大学客員研究員を経て平成18年8月から現職。
専攻:神経科学。
|
 |
科学技術振興機構研究員 加 藤 利佳子 KATO, Rikako
茨城大学理学部卒,筑波大学医学系研究科修了,医学博士。生理学研究所 非常勤研究員,フランス College de France研究員を経て平成17年9月より現職。
専攻:神経生理学。
|
 |
研究員 梅 田 達 也 UMEDA, Tatsuya
東京大学理学部卒,東京大学大学院理学系研究科修士課程修了,東京医科歯科大学大学院医歯学総合研究科修了,医学博士。COE拠点形成特別研究員を経て平成19年4月から現職。
専攻:神経生理学。
|
 |
科学技術振興機構研究員 池 田 琢 朗 IKEDA, Takuro
東京大学理学部卒,東京大学大学院理学系研究科修士課程修了,東京大学大学院医学系研究科博士課程修了,医学博士。科学技術振興機構研究員(さきがけ)を経て平成17年7月から現職。
専攻:神経生理学,認知神経科学。
|
 |
研究員 金 祉 希 KIM, Gee Hee
早稲田大学人間科学部卒,同大学大学院人間科学研究科修士課程修了,同大学院人間科学研究科博士課程満期退学,人間博士。国立障害者リハビリテーションセンター研究所流動研究員を経て平成20年12月から現職。
専攻:神経生理学。
|
 |
研究員 武 井 智 彦 TAKEI, Tomohiko
京都大学総合人間学部卒,同大学大学院人間・環境学研究科博士課程修了,博士(人間・環境学)。平成20年5月から現職。
専攻:神経生理学。
|
 |
研究員 大 屋 知 徹 OYA, Tomomichi
京都大学総合人間学部卒,The University of Queensland, School of Human Movement Studies (Australia) 博士課程修了。(PhD of Neuroscience).平成21年2月から現職。
専攻:神経生理学,運動生理学。
|
発達後期には神経回路機能の再編成がおこる。これは既に機能している神経回路における現象であるため,しばしば行動,感覚やリズムなど個体の脳機能の大きな変化として表現される。
この発達期における神経回路機能の再編成のメカニズムを回路およびシナプスレベルで解明する。
さらに,この変化に対する個体環境/神経活動による制御機構を解明する。
また,外傷や虚血などの障害後には未熟期の回路機能の性質が再現し,回復期には発達と同じような回路機能の変化が再現される可能性について,種々の穿孔パッチクランプ法を始めとする電気生理学的手法や分子生物学的手法で解析を行なっている。神経回路発達に対して特に重要であり,それ自体に著明な変化がおこるGABA作動性回路について,発達制御機構を含めて解明を進めている。
また,生体における発達・回復期における再編機構の観察のため,in vivo多光子顕微鏡による神経回路の可視化を行っており,現在,大脳皮質全層における神経回路の可視化技術の確立を行っている。
(1)成熟における代表的抑制性伝達物質であるGABA/グリシンは,未熟期にはシナプス後細胞に脱分極を引き起こし,直接および間接的に興奮性作用を示す。この発達に伴うGABA/グリシンの興奮性から抑制性極作用のスイッチに関して,細胞内クロールイオンくみ出し分子であるカリウム-クロール共役分子(KCC2)を中心に,細胞内クロールイオン調節機構の発達および障害時における変化,およびその制御機構について検討を進めている。
(2)新たに見つかった神経回路の発達再編である「神経伝達物質自体が単一神経終末内でGABAからグリシンへスイッチする」メカニズムについて,神経終末内関連分子,制御因子や回路活動との関連などを多角的に解析する。また,なぜ未熟期にはGABAが重要であるのかを検討する。
(3)神経成長因子によるGABA受容体機能の修飾作用の発達変化を大脳皮質感覚野において検討しており,感覚入力による制御について検討している。脳由来神経成長因子(BDNF)は視覚野細胞において未熟期では膜表面GABAA受容体の急速な減少を引き起こすが,成熟期には増加へとスイッチすることを明らかにした。この変化と発達期における感覚入力との関連,および細胞内メカニズムの解明を進めている。
(4)障害後におけるグルタミン酸受容体やGABA受容体機能の可塑的変化について,NMDA受容体未熟機能再現やGABAの興奮性作用の再獲得のメカニズムについて,多角的に解析を加えている。
(5)2光子顕微鏡による観察法により生体マウス大脳皮質全層の可視化を行っている。この技術により発達および障害後の神経細胞樹状突起やスパインの変化,およびグリアの動きの観察を試みている。ミクログリアのシナプス監視のメカニズムと障害時における変化についてマウス生体脳で観察するとともに,シナプスのリモディングへの関与について研究を行っている。
また,慢性疼痛時における大脳皮質感覚野の細胞活動変化をカルシウムイメージングを用いて,また,スパインの動的変化も検討している。
職 員
|
 |
教 授 鍋 倉 淳 一 Nabekura, Junichi
九州大学医学部卒,医学博士,東北大学医学部助手,秋田大学医学部助教授,九州大学医学研究院助教授を経て,平成15年11月から現職。
専攻:神経生理学,発達生理学。
|
 |
准教授 石 橋 仁 ISHIBASHI, Hitoshi
九州大学薬学部卒,同大学院修士課程修了,九州大学大学院医学系研究科博士課程修了,医学博士,熊本大学薬学部講師,九州大学医学系研究科講師を経て,平成19年1月より現職。
専攻:神経生理学。
|
 |
特任助教 渡 部 美 穂 WATANABE, Miho
早稲田大学人間科学部卒,同大学院修士課程修了。日本医科大学大学院医学研究科修了。博士(医学)。生理学研究所非常勤研究員を経て,平成18年4月から現職。
専攻:神経科学。
|
 |
研究員 江 藤 圭 Eto, Kei
静岡県立大学薬学部卒,九州大学大学院薬学府修士課程修了,同大学院博士課程修了,薬学博士。平成21年4月より現職。
専攻:神経科学。
|
 |
研究員 金 善 光 KIM, Sun Kwang
慶熙大學韓醫學科卒,同大學院博士課程修了,韓醫學博士。現同大學鍼灸經絡科學硏究所硏究員。現JSPS外國人特別硏究員。
專攻:神經科學。
|
ヒトをはじめとする動物生体は,内的ならびに外的環境の変化に即応しながらも体内の内部環境をできるだけ一定に保とうとする機構を備えており,広くホメオスタシス(恒常性維持機構)として知られている。とりわけ視床下部は,ホメオスタシスの調節系である自律神経系,内分泌系,免疫系をとりまとめる高位中枢として,個体の生命保持ならびに系統維持のための基本的な諸活動を調整する働きを営んでいる。本研究部門では,ホメオスタシスの中でも,特に,摂食行動とエネルギー消費機構からなる生体のエネルギーバランスに注目し,視床下部が生体のエネルギーバランスに対してどのような調節作用を営んでいるかを明らかにすると共に,その破綻が肥満や糖尿病の発症とどう関わるかを解明することを目指している。主たる研究課題は以下の通りである。
(1)視床下部における摂食行動の調節機構。
(2)糖・脂質代謝に及ぼす視床下部-交感神経系の調節機構。
(3)レプチンなどアディポカインの細胞内シグナル伝達機構。
(4)AMPKとAMPKファミリーの代謝への調節作用と病態との関連
|
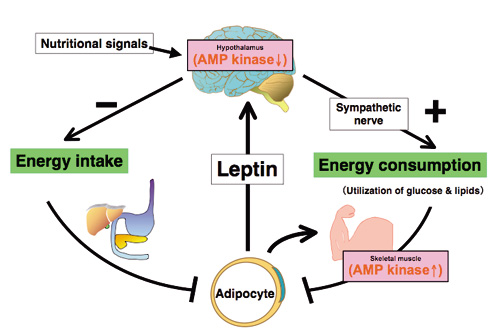 |
図1:レプチンは視床下部と骨格筋のAMPキナーゼ(AMPK)を相反的に調節することによって生体エネルギー代謝を調節する。
レプチンは,骨格筋のレプチン受容体Ob-Rbを介して直接的に,並びに視床下部—交感神経系を介して間接的に骨格筋のAMPKを活性化し,脂肪酸酸化を促進する。またレプチンは,視床下部AMPK活性を逆に抑制することによって摂食抑制作用を引き起こす。AMPK活性に対するこのような相反的な調節作用はレプチンによるエネルギー代謝調節作用に必須である。レプチンが視床下部と骨格筋においてどのようにAMPK活性を相反的に調節するかは,重要な研究課題の一つである。
|
|
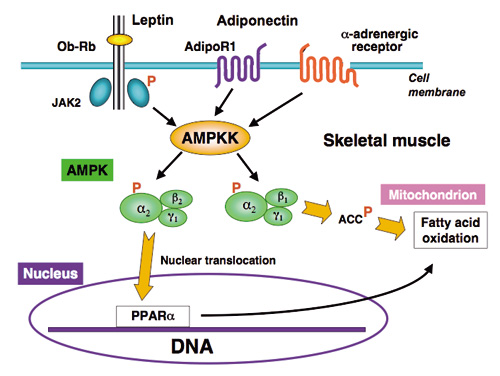 |
図2:骨格筋におけるAMPKの脂肪酸酸化調節作用
レプチン,アディポネクチン及びaアドレナリン作動薬はAMPKKを介して骨格筋a2AMPKを活性化する。a2AMPKのうちb2調節サブユニットを持つAMPKは核に移行してPPARaなどの遺伝子発現を促進,PPARaは脂肪酸酸化関連遺伝子の発現を高める。一方,b1調節サブユニットを持つa2AMPKは細胞質に留まり,ACC (acetyl-CoA carboxylase) 活性を抑制することによってミトコンドリアでの脂肪酸酸化を促進する。
|
|
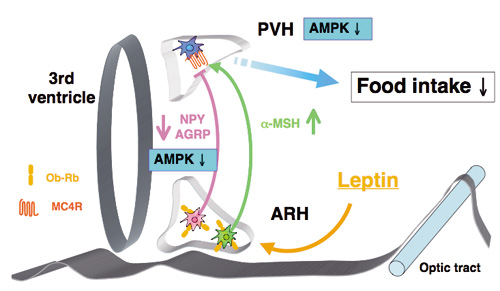 |
図3:視床下部AMPKによる摂食調節作用
弓状核(ARH)には摂食を促進するNPY/AGRPニューロンと摂食を抑制するa-MSHニューロンが存在し,室傍核のメラノコルチン4受容体(MC4R)を拮抗的に調節している。レプチンは,a-MSHニューロンを活性化する一方,AMPK活性を抑制することによってNPY/AGRPニューロンを抑制し,その結果,MC4Rを活性化して摂食を抑制する。活性化したMC4RはさらにPVHのAMPK活性を抑制することによって摂食抑制作用を引き起こす。最近,PVHのAMPKが摂食量だけでなく食餌嗜好性を調節することが明らかとなった。
|
職 員
 |
教 授 箕 越 靖 彦 MINOKOSHI, Yasuhiko
愛媛大学医学部卒,愛媛大学大学院医学研究科博士過程終了,医学博士。同大学医学部助手,講師,助教授,ハーバード大学医学部Lecturerを経て平成15年11月から現職。
専攻:代謝・内分泌学。
|
 |
助 教 志 内 哲 也 SHIUCHI, Tetsuya
徳島大学総合科学部卒,同大学院人間自然環境研究科修士課程修了,愛媛大学大学院医学系研究科博士課程修了,医学博士。愛媛大学医学部助手,平成16年3月より現職。
専攻:代謝・内分泌学,運動生理生化学。
|
 |
助 教 岡 本 士 毅 OKAMOTO, Shiki
北海道大学獣医学部卒,同大学院獣医学研究科博士課程修了,獣医学博士。日本学術振興会特別研究員,東京都臨床医学総合研究所常勤流動研究員を経て平成16年4月から現職。
専攻:神経免疫学,幹細胞生物学。
|
 |
研究員 李 順 姫 LEE, Suni
東京農工大東京農工大学農学部卒,東京農工大学大学院修士課程修了,東京農工大学連合農学研究科博士課程修了,農学博士。東京都老人総合研究所非常勤研究員,ケースウェスタンリザーブ大学およびマイアミ大学において博士研究員を経て平成17年4月より現職。
専攻:神経細胞学,神経科学。
|
|