生体情報研究系
概要
分子生物学的研究により神経系の機能素子やシナプスの研究が飛躍的に進む一方,脳機能イメージングの進歩により大脳皮質等の機能局在の研究が進んでいる。しかしミクロとマクロのレベルをつなぐ神経回路でどのように情報が処理されているかについては未解明な事が多く残されている。本研究系では,脳における情報処理機構をトップダウンとボトムアップの両面から研究している。
感覚認知情報研究部門
感覚認知情報部門は視知覚および視覚認知の神経機構を研究対象としている。主にサルの視覚野からニューロン活動を記録し,ニューロンの刺激選択性や,異なる種類の刺激への反応の分布を調べることにより,視覚情報の脳内表現を明らかにすることを試みると共に,さまざまな行動課題時のニューロン活動を分析することにより,それらの視覚情報が知覚や行動にどのように関係しているかを調べている。また最近無麻酔のサルの機能的磁気共鳴画像法(fMRI)による視覚関連脳活動の解析を進めている。具体的な課題としては
(1)物体の表面の属性(色や明るさ)が大脳皮質でどのように表現されているか,
(2)それらの情報がどのように知覚や行動に関係しているか,
(3)視野の離れた場所に存在する要素刺激を統合して一つの物体として認知する仕組み,
(4)さまざまな向きの局所の輪郭の情報がどのように組み合わされて図形パターンが表現されるか,
といった問題に関して実験を行っている。
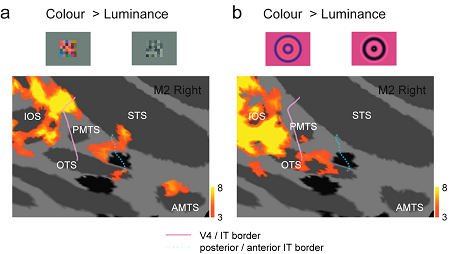
サル下側頭皮質(IT)は色情報処理において重要な役割を果たしている脳領域である。本研究ではサルITにおいて色情報がどのように分布しているかを機能的MRIを用いて明らかにした。図は二種類の視覚刺激 (a: モンドリアン刺激,b:グレーティング刺激)を用いて得られた色選択的活動を示したものである。色選択的活動はIT前部及び後部のそれぞれ数mmの小領域に局在していた。IT前部の活動はモンドリアン刺激によってのみ観察され,IT後部の活動は両刺激によって観察されたがその位置は異なっていた。これらの結果は,ITには色選択性の高い小領域が複数存在し,それらは刺激選択性において異なっていることを示している。
職 員

静岡大学理学部卒,大阪大学大学院基礎工学研究科博士課程修了,工学博士。弘前大学医学部助手,同講師,米国NIH客員研究員,電子技術総合研究所主任研究官を経て平成6年10月から教授(併任),平成7年4月から現職。
専攻:神経生理学。

大阪大学基礎工学部卒,同大学大学院基礎工学研究科博士課程修了,工学博士。理化学研究所フロンティア研究員,米国ロックフェラー大学博士研究員を経て平成10年1月から現職。
専攻:神経生理学。

京都大学工学部卒,同大学大学院人間・環境学研究科博士課程修了,博士(人間・環境学)。(株)国際電気通信基礎技術研究所研究員を経て平成15年9月から現職。
専攻:視覚心理物理学。
神経シグナル研究部門
神経シグナル部門では,神経細胞間および局所神経回路を形成する細胞集団における情報処理のメカニズムを,主に電気生理学的な立場から解析している。また,分子の異常と個体の異常を結びつけるひとつの手段として,自然発症の遺伝子変異もしくは遺伝子改変モデル動物などを用い,複雑な生体システムにおける分子の機能を明らかにしてきている。実験手法としては脳のスライスおよびin vivo パッチクランプ法を用いて,神経回路の機能を系統的に検討している。またカルシウム・カルモデュリン依存性キナーゼIIの遺伝子改変マウスの機能解析を行っている。その他に,分子・細胞レベルからの神経回路理解に向けて,コンピュータを組み込んだ実験(ダイナミッククランプ法)や計算論的なアプローチなども導入しつつある。
主に現在行っている研究は以下のとおりである。
(1)電位依存性カルシウムチャネルの異常により起こる神経疾患の病態解明
本チャネルの異常により,ヒト,マウスで小脳失調症やてんかんなどの神経疾患が起こることが知られている。しかし変異がいかに神経疾患を起こすかに関してはほとんど知見がない。われわれはいろいろな測定方法をあわせて用い,単一の分子の異常が脳機能にどのような影響を与えるかを検討している。
カルシウムチャネルに変異があるてんかんモデルマウスのtottering マウスでは,視床から大脳皮質へのフィードフォワード抑制が顕著に障害されていることを明らかにした(図1)。
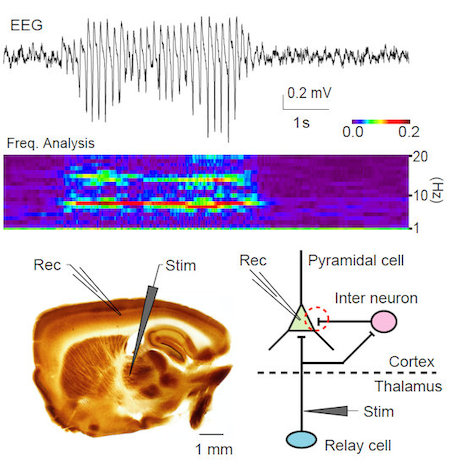
図1.Totteringマウスの欠神発作の脳波(上)。この脳波を周波数解析すると発作中は6 - 7 Hzの周波数成分の増強が見られる(中)。大脳皮質と視床を結ぶ神経線維を保った脳スライス標本(下左)。Totteringマウスでは,視床から大脳皮質細胞への2シナプス性抑制性入力が低下していた(下右,赤破線部)。
(2)In vivo パッチクランプ法を用いた痛覚抑制機構とその異常の解明
独自に開発したin vivo パッチクランプ法を用い,感覚情報,特に痛みや痒みの伝達と,未だ不明なことの多いその抑制機構を解明している(図2)。痛みや痒み情報の中枢への入り口である脊髄後角細胞から記録を行い,生理的な刺激によって誘起される興奮性および抑制性シナプス応答を詳細に解析し,脊髄内における抑制回路を同定している。また,脳幹青斑核からのin vivo パッチクランプ法の開発に成功し,脊髄へ下行性に投射するノルアドレナリン神経を介した抑制機構,さらに,これら内因性抑制系を対象に鎮痛薬や抗掻痒薬の評価を行っている。また,末梢神経(AδやC線維)におけるNaチャネルの解析や行動薬理学的,免疫組織学的解析を併せて行って統合的な解析を行うとともに,神経因性やガン性疼痛の発症メカニズムの解明も行っている。
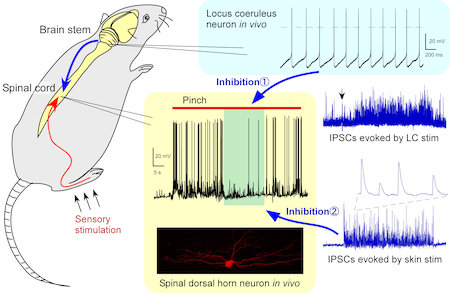
図2.脳幹および脊髄からのin vivo パッチクランプ記録法。痛覚刺激によって発生した脊髄後角細胞の活動電位の抑制機構。①青斑核から下行性にノルアドレナリン神経を介して,②皮膚への触刺激によって,GABA作動性抑制性シナプス応答(IPSC)が賦活化された。
(3)異種シナプス間拡散性クロストークの分子的基盤
神経細胞は,シナプスを介して単一方向のみならず逆行性や拡散性にも情報を伝達する可能性が指摘されている。私たちは,①脳幹の下オリーブ核から小脳プルキンエ細胞へ投射する登上線維の反復刺激に伴い,籠細胞から同じプルキンエ細胞に入力するGABA作動性シナプス伝達が一過性に抑制されることを発見し,②この異種シナプス抑制は,登上線維終末から拡散したグルタミン酸が籠細胞の軸索終末に存在するAMPA受容体を活性化することにより惹起されていることを報告した。現在,この“拡散というユニークな情報伝達機構”が,神経細胞やグリア細胞のグルタミン酸輸送体(EAAT4, GLAST, GLT-1)によって制御されるメカニズムを追究している(図3)。
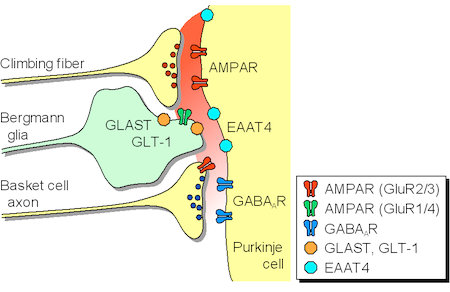
図3.登上線維から放出されたグルタミン酸は,拡散して籠細胞の前シナプス性AMPA受容体(GluR2/3)に作用することにより,籠細胞のGABA放出を阻害する。グルタミン酸のシナプス外拡散は,グルタミン酸輸送体(EAAT4/GLAST/GLT-1)によって厳密に制御されている。
(4)蛋白質リン酸化によるシナプス可塑性,学習・記憶の制御
Ca2+/カルモジュリン依存性プロテインキナーゼIIα (CaMKIIα) は,特に海馬に多く存在する蛋白質リン酸化酵素で,脳機能の調節,中でもシナプス可塑性,学習・記憶の制御に重要な役割を果たすと考えられている。我々は,CaMKIIαのキナーゼ活性をなくした不活性型ノックインマウス [CaMKIIα (K42R)]を作製し,その解析を行うことにより,CaMKIIαによる蛋白質リン酸化が脳機能に果たす役割を解析している。CaMKIIα (K42R)ノックインマウスでは,CaMKIIαの蛋白発現や刺激によるシナプス部への移行が保たれているにも拘わらず,海馬シナプス可塑性,海馬依存性の学習・記憶が強く障害されていた(図4)。現在,これら障害に関わる分子メカニズムの解明と,海馬以外が関与する学習・記憶への影響について研究を重ねている。
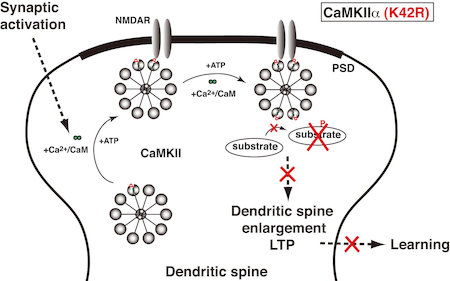
図4.CaMKIIαキナーゼ活性による海馬シナプス可塑性,学習・記憶の制御。不活性型CaMKIIα(K42R)ノックインマウスでは,シナプスの活性化の際に,CaMKIIαのシナプス部への移行は起こるが,そこで基質蛋白質をリン酸化できないために,樹状突起スパインの増大やシナプスの長期増強が起こらず,また,学習・記憶も成立しない。
職 員

京都大学医学部卒,医学博士。国立療養所宇多野病院医師,京都大学医学部助手,講師,助教授,マックス・プランク医学研究所研究員を経て,1995年4月から現職。
専攻:神経生理学。

九州工業大学大学院情報工学研究科博士課程修了,情報工学博士。佐賀医科大学助手,九州大学医学研究院助手,助教を経て,2009年2月から現職。
専攻:神経生理学。

京都大学大学院医学研究科博士課程修了,医学博士。京都大学医学部助手,ロックフェラー大学研究員を経て,1991年9月より現職。
専攻:生化学,神経化学。

名古屋大学大学院理学研究科博士課程修了,博士(理学)。三菱化学生命科学研究所博士研究員,科学技術振興事業団CREST研究員を経て,2002年9月より現職。
専攻:神経生理学,生化学。

総合研究大学院大学生命科学研究科博士課程修了,博士(理学)。2009年4月より現職。
専攻:神経生理学。

九州大学大学院医学系学府博士課程単位取得退学。2010年4月より現職。
専攻:神経生理学。
神経分化研究部門
岡崎統合バイオサイエンスセンター
時系列生命現象研究領域
兼務
(1)大脳皮質視覚野の神経回路の解析
大脳皮質の中でも一次視覚野は,個々の細胞の視覚刺激に対する反応選択性やコラム構造が明確であるため,脳機能とシナプス・神経回路の関係を直接対応付けて解析するのに適した脳領域であると考えられる。そこで,我々は,マウスやラットの大脳皮質視覚野から作成したスライス標本を用い,複数の細胞からの同時ホールセル記録法,ケージドグルタミン酸によるレーザースキャン局所刺激法等を組み合わせて,その神経回路を解析し,以下のような成果を上げている。1)2/3層錐体細胞へ入力を送る細胞の空間分布を調べた結果,興奮性結合している錐体細胞ペアは,別の2/3層錐体細胞や4層細胞からも高い割合で共通の興奮性入力を受けており,非常に微細なスケールの神経回路網を形成している結果を得た。2)興奮性錐体細胞の単発発火によって,近傍の別の錐体細胞に非常に短潜時の抑制性反応が生じることを見出した。2/3層において解析した結果,錐体細胞の軸索が抑制性細胞の樹状突起・細胞体を介さずに,直接抑制性細胞の軸索終末を活性化し,伝達物質を放出させるという全く新しいタイプの回路による抑制反応であることが判明した。現在,このようなシナプス・神経回路の特性や制御機構の解析をさらに進めると共に,視覚情報処理における役割を明らかにするために,遺伝子工学的手法を併用した解析や,麻酔動物を用いた視覚生理実験を行っている。
(2)大脳皮質視覚野の経験依存的発達メカニズム
個々の一次視覚野ニューロンは,特定の視覚刺激に選択的に反応するが,この反応選択性を作り出す機能的神経回路の形成には,遺伝的に定められた神経結合形成と,生後の視覚体験に依存した可塑的調整の2つの段階がある。このため,生まれた時点では未熟な選択性は,発達に伴い成熟する。我々は,視覚反応の経験依存的な発達機構を明らかにするために,様々な発達段階にある動物および暗室飼育等により視覚入力を操作した動物,分子生物学的手法により特定の分子の発現を制御した動物の視覚野を用いて,その神経回路やシナプス可塑性および視覚反応可塑性を調べている。
(3)運動機能の基盤となる神経回路の形成
神経回路は,転写因子の発現と活動依存的な修飾機構により規定される個々のニューロンにより構成される。特定のニューロンは神経回路機能に見合った特性(イオンチャネルによる膜興奮性や伝達物質の種類)を獲得する。発生過程において個々のニューロンが生まれ神経機構が成立するメカニズムを,トランスジェニックゼブラフィッシュなどを用いて解析している(東島准教授ほか)。
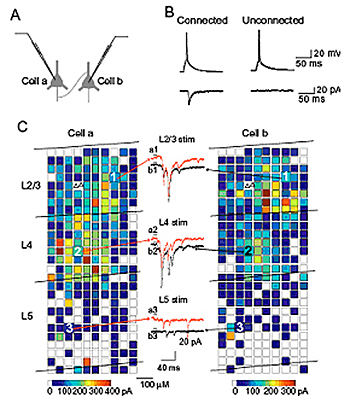
図1 ケージドグルタミン酸を用いた光刺激法による局所神経回路の解析
シナプス結合がみられた2個の2/3層錐体細胞への興奮性入力のマップを左右にそれぞれ示す。2つの△は記録したニューロンペアの位置を示す。中央は矢印で示した場所の刺激に伴い2個の細胞から同時に記録されたEPSCの例。興奮性結合しているペアにおいては,2/3層刺激および4層刺激によるEPSCのタイミングがかなり一致している。マップ上の各□で示した部位をそれぞれ刺激したときに生じたEPSCの大きさを擬似カラー表示している。
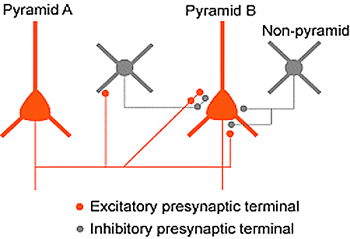
図2 抑制性細胞の細胞体・樹状突起ドメインをバイパスする錐体細胞間の抑制性伝達
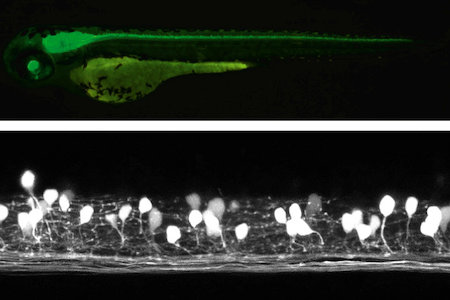
図3 生きたままニューロンを蛍光タンパクの発現によって可視化したトランスジェニックゼブラフィッシュ。上図は通常の蛍光写真。下図は共焦点顕微鏡画像。
職 員

大阪府立大学卒,大阪大学大学院医学研究科修士課程終了,同博士課程修了,医学博士,日本学術振興会特別研究員(大阪バイオサイエンス研究所),名古屋大学環境医学研究所助手,ソーク生物学研究所研究員,名古屋大学環境医学研究所准教授を経て平成21年2月から現職。
専攻:神経生理学。

東京大学理学部生物化学科卒,同大学院博士課程修了,理学博士。基礎生物学研究所助手,科学技術振興事業団さきがけ研究専任研究員,ニューヨーク州立大学ストーニーブルック校客員研究員を経て平成15年11月から現職。
専攻:神経生理学,発生神経科学。

京都大学理学部卒,同大学院理学研究科博士課程修了,博士(理学)。京都大学霊長類研究所博士研究員,ソーク研究所博士研究員を経て,平成21年3月より現職。
専攻:神経生理学,ウイルス学。

総合研究大学院大学生命科学研究科卒,理学博士。平成18年4月から現職。
専攻:神経解剖学。

埼玉大学卒,東京大学理学系研究科修了,理学博士,生理研研究員を経て平成19年4月から現職。
専攻:発生生物学。

大阪大学基礎工学部卒,大阪大学生命機能研究科博士課程終了,理学博士。日本学術振興会特別研究員を経て,平成21年4月から現職。
専攻:神経生理学。

早稲田大学教育学部卒,総合研究大学院大学生命科学研究科修了,理学博士。平成22年4月から現職。
専攻:神経生理学。