大脳皮質機能研究系
概要
大脳皮質機能研究系は脳形態解析研究部門,大脳神経回路論研究部門,心理生理学研究部門の専任3部門からなり,分子・神経細胞レベルから神経回路・個体脳レベルへの統合を目指した研究を活発に進めている。脳形態解析研究部門では,特にグルタミン酸受容体やイオンチャネルの局在と神経伝達調節における機能,記憶の長期定着メカニズム,脳の左右非対称性について研究を行っている。大脳神経回路論研究部門では,新皮質を構成するニューロンを形態・分子発現・電気的性質・皮質外投射などから同定した上で,神経細胞タイプごとのシナプス結合パターンや in vivoにおける発火特性などを生理学と形態学の手法を組み合わせて調べている。心理生理学研究部門では,認知,記憶,思考,行動,情動,感性などに関連する脳活動に注目して,主として機能的MRIを用いてヒトを対象とした実験的研究を推進している。
脳形態解析研究部門
脳における情報のやりとりは,神経細胞膜上に存在する,伝達物質受容体やチャネル,トランスポーターなどの分子によって担われ,制御されている。本部門ではこれらの分子の局在関係や動態を詳細に観察し,シナプス,神経回路,システム,個体行動の各レベルにおける,機能分子の役割を分子生物学的,形態学的および生理学的方法を総合して解析する。特に,各レベルや方法論のギャップを埋めることによって,脳の統合機能の独創的な理解を目指す。最近,凍結割断レプリカを用いた標識法で生体内分子の局在を1分子レベルで同定する新たな方法論を開発し,記憶学習に伴う生体内分子動態を電気生理学や行動学的方法と合わせて解析している。
(1)グルタミン酸受容体およびGABA受容体の脳における電子顕微鏡的局在を,特にシナプスとの位置関係や,受容体刺激によって制御される各種チャネル分子との共存に注目して,解析する。特に,凍結割断レプリカ免疫標識法を用いて,分子の数やそれらの共存関係を電子顕微鏡レベルで解析し(図1),記憶,学習の基礎となる,シナプスの可塑的変化に伴う分子の動きを解析する。これらの結果と,個体の学習行動や電気生理学的実験,培養細胞などのモデル系を用いた機能分子の局在可視化を組み合わせ,機能分子の局在制御メカニズムと生理的意義を探る。また,最近発見した海馬シナプスにおけるNMDA受容体局在の左右差(図2)について,その意義と非対称性の生成メカニズムを探る。
(2)シナプス局在性細胞接着因子Neurexin/Neuroligin の神経回路網形成における役割と,自閉症をはじめとする精神・神経疾患との関連について研究する。主に,これらの遺伝子改変マウスを作出し,それらの興奮性および抑制性シナプスに対する影響を,生化学,電気生理学,電子顕微鏡などの手法を用いて解析するとともに,社会的相互作用,空間学習記憶などの行動を,疾患との関連から解析する(図3)。
(3)シナプス−グリア複合環境の変化が,伝達物質濃度の時空間特性にどう影響するのか調べる。これまで,シナプス前細胞からグリア細胞に向けて異所性のシナプス小胞放出があり,これがニューロン−グリア間の素早い情報伝達を担っていることを示してきた(図4)。この情報伝達によってグリア細胞の形態が制御されている可能性を,二光子励起イメージングによって解析する。グリア細胞によるシナプスの包囲率の相違が,シナプス伝達にどんな影響を与えるのかを,電気生理学・電子顕微鏡法も組み合わせて解明する。
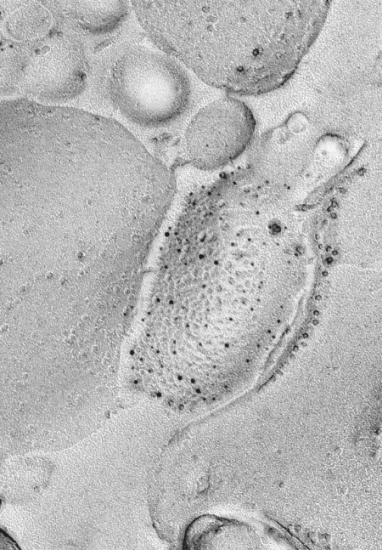
図1:AMPA型(5nm金標識)およびGluRd2型(10nm金標識)グルタミン酸受容体の小脳における共存
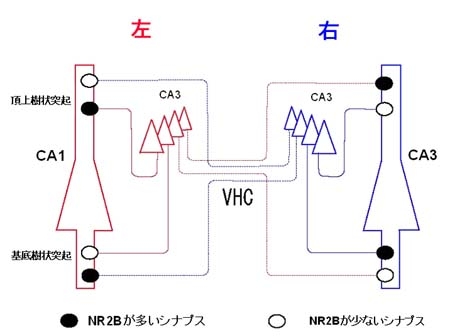
図2:NMDA受容体配置の左右非対称性
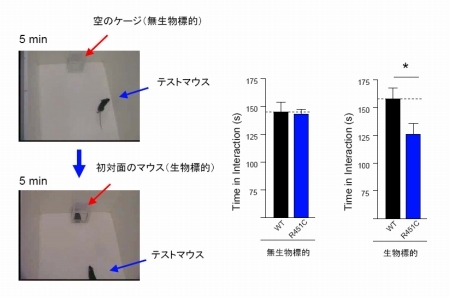
図3:Neuroligin-3 のR451Cノックインマウス では,社会行動の異常が見られた。
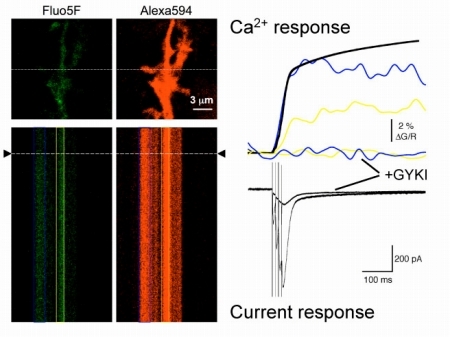
図4:シナプス刺激によるグリア細胞のCa2+上昇と電流応答の同時記録
職 員

京都大学医学部卒。医学博士。京都大学附属病院神経内科にて研修後,京都大学医学部助手を経て,平成10年12月から現職。
専攻:神経解剖学,分子神経科学。

筑波大学医学専門学群卒。大阪大学大学院医学研究科修了,医学博士。ハワードヒューズ研究員,テキサス大学研究員,Instructor,スタンフォード大学医学部Research Associateを経て,平成21年4月から現職。
専攻:神経解剖学,分子神経科学。

横浜市立大学文理学部卒。理学博士。三菱化学生命科学研究所特別研究員を経て,平成13年10月から現職。
専攻:内分泌学,分子神経科学。

東京大学文学部卒。東京大学大学院人文社会系研究科博士課程修了,心理学博士。日本学術振興会海外特別研究員,上原記念生命科学財団リサーチフェローシップ,Oregon Health & Science Universityポスドク研究員を経て,平成18年2月から現職。
専攻:神経生理学。

日本女子大学家政理Ⅱ学科卒。理学博士。三井化学総合研究所,日本女子大学非常勤講師,Colorado State University,研究員,Research Assistant Professor を経て,平成19年4月から現職。
専攻:神経超微構造学。

大阪大学理学部卒。大阪大学大学院生命機能研究科修了,理学博士。日本学術振興会特別研究員を経て,平成22年4月より現職。

九州大学理学部卒。九州大学大学院医学系研究科博士課程修了,理学博士。九州大学大学院学術研究員,学振特別研究員を経て,平成20年4月から現職。
専攻:神経生理学。

HARADA, Harumi
秋田大学教育文化学部卒,筑波大学大学院生命環境科学研究科博士課程修了,博士(環境学)。平成22年4月から現職。
専攻:分子神経科学。

第四軍医大学医学部卒。第四軍医大学(西安)博士課程修了,医学博士。第四軍医大学解剖学部准教授。平成20年6月から現職。
専攻:神経解剖学。
大脳神経回路論研究部門
大脳新皮質は形や働きが異なる極めて多様なニューロンからできているが,これらの配置や結合の規則はあまり分かっていない。大脳の領野ごとの機能分化は次第に解明されつつあるが,より微小な単位である皮質局所回路の本質的な動作原理は殆ど明らかにされていない。本研究室の目標は,大脳システムにおける新皮質局回路の構築原理を理解することである。
そのために,現在は齧歯類の1次・2次運動野に焦点をあて,構成ニューロンタイプを皮質外投射・電気的性質・形態・分子発現など多方面から同定し,前頭皮質領域分化を考慮にいれながら,神経細胞タイプごとのシナプス結合パターンや in vivo発火特性等を生理学と形態学の手法を組み合わせて調べている。新皮質局所回路と大脳システム回路を統合的に解析することで,ニューロンタイプの機能分担や層構造の役割,さらに前頭皮質から他の皮質・視床・基底核・小脳などへの多様な投射や,大脳の局所・大域クロックの機能的意味を探求していこうと考えている。
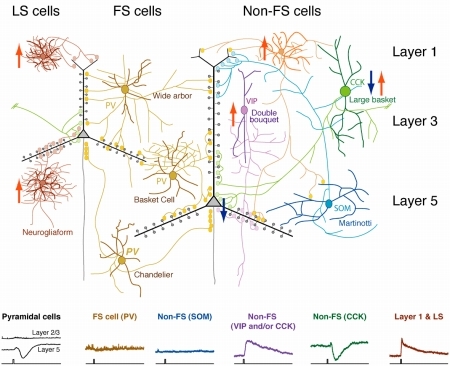
図1 アセチルコリンの大脳皮質ニューロンサブタイプに対する一過性作用。上図:大脳皮質ニューロンタイプと,アセチルコリンによる興奮性効果(上向き赤矢印)と抑制性効果(下向き青矢印)。下図:アセチルコリン短時間投与に対する典型的な応答。持続的投与による結果と合わせると,アセチルコリンは抑制性ニューロンに対してニコチン受容体による脱分極・ムスカリン受容体による過分極・ムスカリン受容体による緩徐な持続的脱分極がサブタイプごとに異なる組み合わせで起きることがわかった。
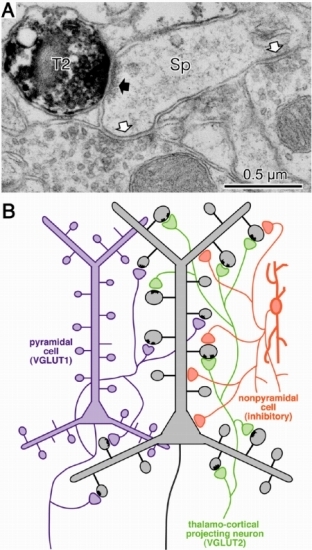
図2 大脳皮質の錐体細胞の棘突起に入力する視床からの神経終末の一部は,皮質の非錐体細胞由来の抑制性神経終末によって直接抑制作用を受ける。A:棘突起(SP)にVGLUT2陽性神経終末(視床由来,黒矢印)が入力しているが,その同じ棘突起には抑制性作用を持つ対称型のシナプスを持つ神経終末2つ(白矢印)が入力している。B:簡単な概念図。視床からの神経終末(緑色)の約1割程度は,抑制性神経終末(赤色)が入力する棘突起を神経支配する。
職 員

東京大学医学部卒,同大学院医学系研究科博士課程修了,医学博士。生理学研究所助手,テネシー大学研究員,理化学研究所フロンティア研究員,チームリーダーを経て,平成11年1月から現職。
専攻:神経科学。

大阪府立大学総合科学部卒,大阪大学大学院医学研究科修士課程修了,同博士課程修了,医学博士。日本学術振興会特別研究員,テネシー大学研究員,香川医科大学助手,ブリティッシュコロンビア大学研究員,理化学研究所基礎科学特別研究員,フロンティア研究員を経て,平成13年10月から現職。
専攻:神経解剖学,神経科学。

大阪大学基礎工学部卒,大阪大学大学院基礎工学研究科修士課程修了,大阪大学大学院工学研究科博士課程修了,工学博士。Duke大学研究員を経て平成16年6月から現職。
専攻:神経科学。

東京薬科大学生命科学部卒,大阪大学大学院医学系研究科修士課程修了,総合研究大学院大学生命科学研究科博士課程修了,理学博士。研究員を経て平成18年12月から現職。
専攻:神経科学。

長崎大学薬学部卒,九州大学大学院薬学府修士課程修了,同大学院同学府博士課程修了,薬学博士。平成19年4月から現職。
専攻:神経科学。

京都工芸繊維大学繊維学部高分子学科卒,奈良先端科学技術大学院大学バイオサイエンス研究科修士課程修了,京都大学大学院医学研究科博士課程修了,医学博士。平成20年4月から現職。
専攻:神経科学。

大阪大学基礎工学部卒,総合研究大学院大学生命科学研究科 5年一貫制博士課程修了,理学博士。日本学術振興会特別研究員を経て,平成22年4月から現職。
専攻:神経科学。

総合研究大学院大学生命科学研究科修了,博士(理学)。平成22年4月より現職。
専攻:神経科学。
心理生理学研究部門
認知,記憶,思考,行動,情動,感性などに関連する脳活動を中心に,ヒトを対象とした実験的研究を推進している。脳神経活動に伴う局所的な循環やエネルギー代謝の変化をとらえる脳機能イメーシングと,時間分解能にすぐれた電気生理学的手法を統合的にもちいることにより,高次脳機能を動的かつ大局的に理解することを目指している。特に,機能局在と機能連関のダイナミックな変化を画像化することにより,感覚脱失に伴う神経活動の変化や発達および学習による新たな機能の獲得,さらには社会能力の発達過程など,高次脳機能の可塑性 (=ヒト脳のやわらかさ) のメカニズムに迫ろうとしている。
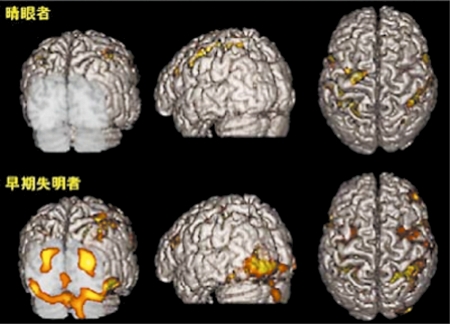
視覚障害者の点字弁別課題における両側一次視覚野の脳賦活
早期視覚障害者における右示指による点字弁別課題中の脳賦活状態を,高分解能MRlに重畳した(下段)。黄色く示した部位で,課題遂行中に統計的有意に血流が増加したことを示している。一方晴眼者(上段)では後頭葉の賦活は全く見られない。視覚障害者では,後頭葉への視覚情報入力が欠損しているにも関わらず,点字読を含む触覚課題によって一次視覚野に劇的な神経活動が生じていることがわかる。幼少時からの視覚脱失により脳の可塑性が発揮されたものと考えられる。
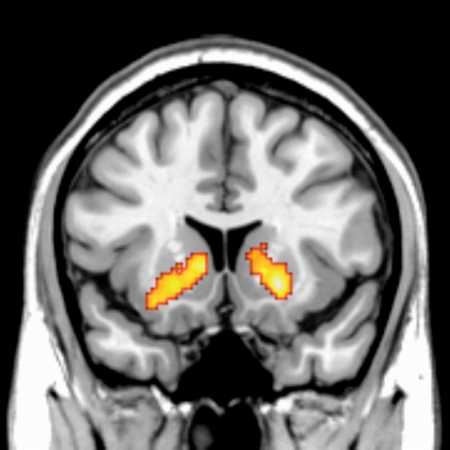
金銭報酬と社会的報酬による基底核の活動
報酬は全ての生物の行動決定に影響を及ぼす要因である。ヒトにおいては食べ物などの基本的報酬の他に,他者からの良い評判・評価というような「社会的報酬」が行動決定に大きな影響を持つということが,社会心理学などの分野の研究から知られている。しかし,今までそのような社会的報酬が,その他の報酬(例えば,食べ物,お金)と同じ脳部位で処理されているのかはわかっていなかった。この研究では,他者からの良い評価を社会的報酬として与えた場合は,金銭報酬を与えた時と同じ報酬系の脳部位が,同じ活動パタンを示すということを見出した。他者からの評判・評価という社会的報酬が,普段の我々の社会的行動に大きな影響を持つことを考えると,この知見は複雑なヒトの社会的行動に対して神経科学的説明を加えるための重要な最初の一歩であると考えられる。
職 員

京都大学医学部卒,同大学院修了,医学博士。米国NIH客員研究員,福井医科大学高エネルギー医学研究センター講師,助教授を経て平成11年1月から現職。
専攻:医療画像,神経科学。

国際基督教大学教養学部卒,大阪大学大学院医学研究科修士課程修了,同博士課程修了,博士(医学)。通信総合研究所柳田結集型特別グループ研究員,生理学研究所科学技術振興機構研究員を経て平成16年12月から現職。
専攻:認知脳科学。

京都大学総合人間学部卒,同大学院人間・環境学研究科修士課程修了,同博士課程修了,博士(人間・環境学)。クィーンズ大学心理学部博士研究員,日本学術振興会海外特別研究員,クィーンズ大学心理学部助手を経て平成20年8月から現職。
専攻:認知脳科学,心理物理学。

上智大学文学部卒業,総合研究大学院大学生命科学研究科博士課程修了,博士(理学)。日本学術振興会特別研究員,日本学術振興会海外特別研究員(米国NIH),ERATO下條潜在脳機能プロジェクト研究員を経て平成22年4月から現職。
専攻:神経科学,認知科学。

東京大学工学部卒業,東京大学大学院工学系研究科修士課程修了。博士(工学)。株式会社日立製作所システム開発研究所研究員,主任研究員を経て平成22年6月から現職。
専攻:認知神経科学。

東京農業大学農学部卒,福井大学大学院教育学研究科修士課程修了,総合研究大学院大学博士課程修了。博士(理学)。平成19年4月から現職。
専攻:神経科学。

京都大学工学部卒,同大学院工学研究科修士課程修了,九州大学医学部卒,同大学院医学系学府博士課程修了。博士(医学)。九州大学病院心療内科助教を経て平成22年4月から現職。
専攻:心身医学,精神神経免疫学。

慶應義塾大学理工学部卒,同大学院基礎理工学研究科修士課程修了。修士(工学),同博士課程修了。博士(理学)。国立身体障害者リハビリテーションセンター研究所流動研究員を経て平成22年4月から現職。
専攻:神経生理,心理生理,音響工学。

東京薬科大学生命科学部卒業,名古屋大学大学院医学系研究科修士課程修了,同博士課程満了,博士(医学)。京都大学大学院情報学研究科・研究員,名古屋大学サイエンスコミュニケーション推進室・サイエンスコミュニケーター等を経て平成22年4月から現職。
専攻:認知神経科学,神経生理学。