| 2. | 上丘局所神経回路(2)中間層ニューロンのバースト活動生成機構の解析(Saito & Isa, J. Neurosci 2003; J. Neurophysiol. 2004) |
|---|---|
| Saito Y, Isa T (2003) Local excitatory network and NMDA receptor activation
generate a synchronous and non-linear bursting command from the superior
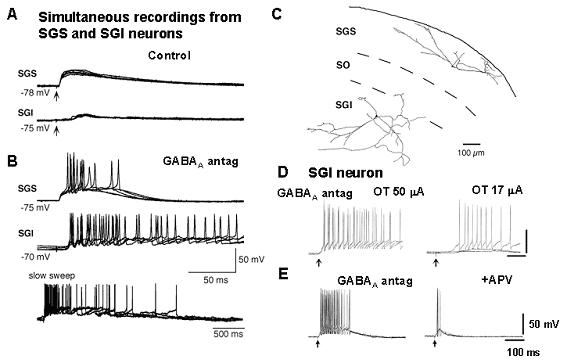
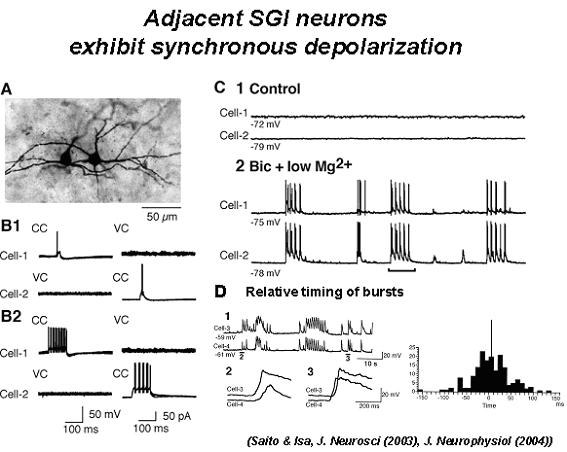
colliculus. Journal of Neuroscience, 23:5854-5864. Saito Y, Isa T (2004) Laminar specific distribution of lateral excitatory connections in the rat superior colliculus. Journal of Neuophysiology, 92:3500-3510. 上丘中間層のニューロンは眼球のサッケード運動などの指向運動の開始に先立って数百ヘルツの高頻度(バースト)発火を生成する。このバースト発火は数ヘルツから数十ヘルツの低頻度発火からいきなり開始されるので、サッケード生成回路のいずこかにサッケードの生成を促進する"decision signal"が「閾値」を超えたことを検出するメカニズムが存在することが示唆されるが、この「閾値検出器」が上丘に存在するのか、より上流の脳領域に存在するかは明らかでない。本研究ではラット上丘スライス標本においてバースト発火の生成機構が上丘に内在するかどうかを検証した。 その結果、上丘スライス標本において中間層ニューロンからホールセル記録を行い、浅層の電気刺激を行い、単シナプス性EPSPを誘発しながら細胞外液に10 μM bicucullineを投与すると数百ミリ秒から数秒持続する脱分極応答と20-30発の活動電位が誘発されること、そしてこの脱分極応答はしばしば全か無(All or none)に誘発されることから、GABA作動性抑制が減弱すると、上丘内の局所神経回路でバースト発火が誘発されるという性質があることが明らかになった。そしてこのバースト発火はAPVの投与で抑制されることからNMDA型グルタミン酸受容体依存性であることが見出された。 そして上丘中間層の近接する2個のニューロンから同時にホールセル記録を行い、細胞外液に10 μM bicucullineを加え、Mg2+濃度を0.1 Mに減らすと、これら2個のニューロンは同時に自発的な脱分極とバースト発火を繰り返すことが見出された。この自発性同期的脱分極はNMDA受容体依存性であり、シナプス入力に対するバースト発火ときわめて類似した性質を有していた。さらにこの同期的脱分極は局所神経回路における興奮性結合に依存したことから、上丘中間層にはGABA作動性抑制が減弱した状況で局所神経回路を構成するニューロンが興奮性結合を開始して同期発火する機構が存在し、それがバースト発火を生成する基本的メカニズムになっていることを示している(以上Saito Y, Isa T, 2003)。以後、bicuculline、及び低Mgの条件で誘発される自発性同期的脱分極を上丘におけるバースト発火生成の基本的メカニズムであると考え、その生成機構を詳細に調べることにした。 次にこの自発性同期的脱分極の起きかたを、浅層ニューロン、視神経層のwide field vertical cellと中間層ニューロンで比較した。すると中間層ニューロンとwide field vertical cellでは大多数のペアで観察されるのに対して浅層ニューロンではごくまれにしか観察されないことから、中間層ニューロンに特有な神経回路機構によるものと考えられる。そして同時記録する2個のニューロンの距離を次第に離していくと、浅層ニューロンで観察される自発性同期的脱分極は両者の距離が100 μmを超えると観察されなくなるのに対し、中間層ニューロンでは距離とともに次第に同期性は減弱し、1ミリ離れたときに同期性が失われることが明らかになった。Wide field vertical cellにおいては数百ミクロンの距離で同期性が失われるというその中間的な性質が見られた。以上の結果から上丘浅層における水平方向の興奮性伝播は狭い範囲に限局しており、浅層が「コラム状」の機能単位から構成されているのに対して中間層は興奮性信号伝達が広範に存在しており、距離とともに興奮性結合が次第に減衰していくという構造になっていることを示している(以上、Saito Y, Isa T, 2004)。 |
|
 |
|
 |
|
| 研究室TOPへ | 生理研のホームページへ |