江橋先生の"トレードマーク"図に想う― 江橋先生6周忌と生誕89周年に寄せて ―
所長 岡田 泰伸
江橋節郎先生がお亡くなりになってこの7月17日でまる5年(6周忌)が過ぎ、8月31日にご生誕89周年を迎えます。江橋先生は1985-91年に生理研所長、1991-93年に岡崎国立共同研究機構長を務められ、2006年に亡くなられるまで名誉教授として岡崎で研究を続けられると共に、生理研や私達後輩のために数々のご尽力をいただきました。江橋先生は、筋肉の弛緩因子は筋小胞体であり、それはアクチン収縮をもたらすCa2+を取り込むことで作用するということを明らかにされ、筋興奮・収縮関連に関する「Ca説」を打ち立てられました。更には、Ca受容蛋白質トロポニンを発見され、トロポニンの筋収縮抑制作用がCa2+結合によって解除されるというCa説の分子メカニズムを明らかにされ、これによって細胞内シグナルとしてのCa2+の役割を世界で初めて明確にされました。それゆえ、私は常々、江橋先生はたとえれば「ノーベル生理学・医学賞」2つ相当のお仕事をされたと申し上げているのです。
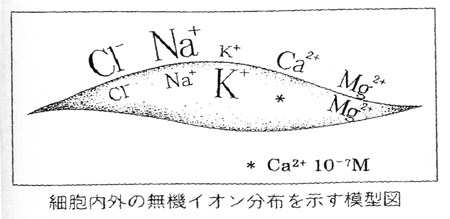
この図は、江橋先生が講演される時にいつも最初に示されるスライドで、いわば江橋先生の“トレードマーク”のような図です。これはもちろん細胞内でCa2+濃度が極端に少ないことを示し、Ca2+の細胞内シグナルとしての前提条件を述べるイントロとして用いられることが多かったのですが、私は他にも多くの含蓄を見出します。現代生理学研究は、ここに示された細胞内外イオン分布差をめぐって展開されて来たようにみえるのです。それは、神経活動電位の発生(脱分極から陽電位へのオーバーシュート)とそれからの回復(再分極化)が、それぞれNa+流入電流とK+流出電流によるというHodgkin-Huxleyの「Na+説」にはじまります。その後、この流れの研究は、京大の沼正作先生のグループを中心にして多くの研究者の努力によって実現された電圧作動性Na+チャネルや電圧依存性K+チャネルの遺伝子クローニングと、Neherらによるパッチクランプ法の開発とそれを駆使しての多くの研究へと発展しました。この研究には、今岡崎で活躍している野田昌晴先生や井本敬二先生、そして久保義弘先生らが大きな寄与をされました。
Ca2+については江橋先生の研究から西塚泰美先生(神戸大)によるCキナーゼ、垣内史郎先生(阪大)によるカルモデュリンの発見をはじめとする細胞内Ca2+シグナル研究の流れと、沼研の田辺勉先生、森泰生先生、竹島浩先生らによる電圧作動性Ca2+チャネルとリアノジン受容体Ca2+放出チャネルや、御子柴克彦先生(当時阪大)によるIP3受容体Ca2+放出チャネルの遺伝子クローニングに至る流れがありました。この前者の流れは、現在のCa2+以外の数多くの細胞内シグナル研究のうねりにつながっています。また後者の流れは、非電圧作動性Ca2+流入路を与えると共に多くの環境因子に対するセンサーとしての働きもするTRPカチオンチャネルの研究という(現在最も高みにある)大波につながっていきます。米国Julius研での最初の哺乳類TRPチャネル発見には、富永真琴先生が大きな寄与をされ、富永先生は現在もこの分野で大活躍されています。
この図は、Cl−も細胞内濃度は細胞外に比べて低いことを示しています。殊に成熟ニューロンでは極めて低濃度に(K+- Cl−コートランスポータ(KCC)の働きによって)維持されており、GABA/グリシン作動性クロライドチャネルとしてのGABAA受容体やグリシン受容体は、Cl−流入電流によって細胞内電位をよりネガティブにすることによって興奮性を抑制する働きをもたらします。これらのGABAA受容体やKCCの研究には、鍋倉淳一先生が大きな寄与をされています。Cl−を分泌する腺細胞においては、細胞内Cl−は細胞外よりは低濃度ではあるが、負の細胞内電位で受動的に分配される濃度(電気化学的平衡濃度)よりは高い濃度にあり、そこに発現しているCFTRクロライドチャネルの開口はCl−流出をもたらすことになります。多くは電気化学的平衡濃度に近い細胞内Cl−濃度を持つその他の殆どの細胞種においてもK+チャネルが同時的に開口してもたらす過分極(細胞内静止電位のよりネガティブ化)によってCl−流出が引き起こされることになり、逆に何らかの原因によって脱分極(細胞内静止電位のよりポジティブ化)がもたらされることによってCl−流入が引き起こされることになります。その結果、(私達が世界で初めて示し、現在広く認められているように)Cl−チャネルは細胞容積調節(とそれが大きく関与する細胞移動や細胞分裂・増殖の調節)や細胞死誘導に不可欠の役割を果たすことになるのです。また、(これも私達が多くの事例と証拠で示したのですが)たとえ細胞内静止電位の変化はなくとも、細胞内に高い濃度で存在するグルタミン酸やATPなどの有機アニオンはクロライドチャネル開口によって細胞外へとそこから流出して、細胞外シグナルをもたらすことになるのです。これらの細胞生死や細胞間シグナリングに深く関与するクロライドチャネルには、その性質から2種類存在していることが判っていますが、いずれも未だ分子同定がなされていません。その遺伝子クローニングが実現されれば、この分野の研究は多くの他の研究分野をまきこんでの一大潮流を生み出していくことになるものと、現在、私達のグループはSabirov客員教授のグループと協同して必死に取り組んでいるところです。
江橋先生は、所長に就任された時に、これからの生理学研究の中心は脳神経研究へと移ると見越されて、そのような舵取りをされました。同時に、高次脳機能の研究の発展には、分子・細胞レベルの一般生理学的研究や生物物理学的研究の支えがなくてはならないとも考えておられ、その後の人事もそのように進められました。そのお考えの基礎には、この“トレードマーク”図があったものと私は推測しています。
来年は、江橋先生生誕90周年を迎える記念の年となります。生理研の研究にも記念すべき年となることを祈念しています。
(“NIPSかわらばん”27号 2011.8.29発行 掲載)