9岡崎統合バイオサイエンスセンター
9.1ナノ形態生理研究部門
1.位相差電子顕微鏡の開発と応用(永山G)
位相板の開発と位相差法の生物応用を行い評価を得た(永山、Danev、大河原、新井、細川:2007年度文部科学大臣表彰)。昨年に引続き、アハラノフーボーム(AB)効果を利用する無損失位相板の開発に注力した。2006年10月から開始したCRESTプロジェクト「ns-nm分解能電子・光子ハイブリッド顕微鏡の開発」の事実上の初年度として、新規電子顕微鏡の要素技術、光電子顕微鏡、電子・光子ハイブリッド顕微鏡などの開発を行った。また、同一試料、同一視野、同一倍率での電顕と光顕の客観的比較を行い、興味ある知見を得た。 数年来幅広く共同研究を行い、種々の生物試料を観察してきたが(Shimada et al, Cell (2007); Yui et al. Langmuir (2007)) 、医学生物学への応用として以下のテーマに絞り込んだ。 i)膜蛋白質の単粒子解析, ii)バクテリアのその場観察と核酸動態, iii)培養細胞のその場観察と細胞内動態(特に細胞骨格)。 i)に関しては、位相差法と従来法の比較のためGro ELを標準蛋白質として解析を行い、位相差法の優位性を示した(Danev & Nagayama, J Stru Biol (2008))。 ii)についてはDNAの局在観察手法(10nm分解能),電子多色イメージング,の開発を始め、シアノバクテリアのDNA局在について新知見を得た(Kaneko et al. J. Plasma and Fusion Res.(2007))。 iii)については、電顕光顕観察用神経細胞系の開発を開始した。また、位相差法を百万ボルト電顕に拡張する試みを韓国の基礎科学研究所との共同研究として開始した。
2.唾液分泌機構(村上G)
傍細胞輸送調節の基盤として存在するタイト結合の変化を凍結割断標本の電子顕微鏡観察を継続した。唾液分泌増強を増強するとされる漢方薬20種類について南京医科大学と共同で摘出灌流顎下腺を用い調査し15種類に唾液腺に直接作用があること、5種類は神経/内分泌などの制御を介すことを示した。増強パターンは3種類に分類され、漢方薬の分類を反映した。現在引き続き増強機序を実験的に検討している。カトリック大学ローマ校との唾液蛋白の共同研究も継続している。
3.質量顕微鏡の開発と多次元メタボローム脳解剖への応用(瀬藤G)
質量顕微鏡は組織切片の観察機能、その組織切片へレーザーを直接照射することによって切片上に存在する生体分子を同定する機能、それらを組み合わせた装置である。現在、生物学的、病理学的なサンプルへの検討に力を入れている。大腸癌組織切片の解析では、癌部位における生体分子の特異的な蓄積が明らかとなった。さらにこの生体分子ついて多段階質量分析を行った結果、Sphingomyelinであることが判明した。この結果は、他の研究によって明らかにされてきた結果を支持すると同時に、我々が開発してきた質量顕微鏡を用いた生体試料からの直接分析方法が、生体上において起こっている現象を正確に捕えることに有用であることを証明した。また、これらの検討と同時に独自に開発してきた質量顕微鏡プロトタイプ機は2号機まで作成され、現在の規格ではMALDIイオン化部を大気圧環境にすることができるようになっている。これによって“生のまま試料を観察する”ことが可能になった。
4.エンドサイトーシス後期の分子過程(大橋G)
細胞外分子取り込みや生物の発生過程において、エンドソーム-ゴルジ細胞内膜系の担う生理機能についての研究を行った。
5.構造変化型ユニバーサル塩基の開発(片岡G)
相対する塩基が特定の塩基でなくともDNA二重鎖の形成を維持する核酸塩基、pyrimido [4,5-d] pyrimidine-2,4,5,7-(1H, 3H, 6H, 8H)-tetraoneを開発した。この人工の核酸塩基は分子内水素移動による迅速なケト-エノール互変異性化と、プリン型-ピリミジン型塩基構造の配 座異性化によって、相対する塩基に呼応して構造変化し、アデニン、グアニン、シトシン、チミンのすべてと塩基対を形成する。概念的に全く新規な核酸塩基であり、様々な生化学ツールとしての利用が期待される。
9.2神経分化研究部門
1.電位感受性酵素の動作原理
Ci-VSP(Ciona voltage-sensing phosphatase)はイオンチャネルの電位センサードメインとホスファターゼドメインを併せ持ち、膜電位依存的に酵素活性を変化させる。これまでイオンチャネルの中にbuilt-inされイオンの透過に特異的に関わると考えられてきた膜電位センサーが、独立した機能ユニットであり、その役割がイオンの透過に留まらず幅広い生理現象に関わる可能性を初めて示した(Murata et al, 2005)。2007年度は、この分子の動作原理の解析を進め、PHドメインGFPを用いたPIP2のイメージングとK チャネルの強制発現による電気生理を並行して行い、脱分極でPIP2のレベルが減少することを明らかにした(Murata & Okamura, J. Physiol.)。また、酵素ドメインの基質特異性をマラカイトグリーンアッセイとTLC解析により行い、これまで明らかになっていたPIP3だけでなくPIP2が脱リン酸化されることが明らかになった(図)。これらの結果から、VSPには多くの電位依存性チャネルと同様に(1)脱分極により下流の分子機能を活性化する、(2)膜電位センサーの動きを翻訳して、下流の活性をチューニングできる、という「電位センサー蛋白」に共通する原理があてはまることが示唆された。
さらに、通常蛋白ドメイン間の相互作用は一方向性ではなく双方向性であることが知られているので、電位センサーの動きが細胞内ドメインの構造変化によりどのように影響を受けるかを検討したところ、活性中心の変異の導入、またはホスファターゼの阻害剤による酵素活性の抑制時に、ゲート電流の動きが有意に速くなるという現象を見出した。このことは電位センサードメインが酵素ドメインと強くカップルして構造変化を生じていることを示唆しているとともに、今後ホスファターゼのダイナミクスを明らかにする端緒になると期待される(Israil ら、Biophysical Society meeting)。
更にVSPがどのような生物現象の文脈での膜電位変化に対応して機能するのか、哺乳類に固有の生理機能の進化とどのような関係があるか、などを明らかにするため、各動物のオルソログ分子の発現様式と分子機能を解析している(村田、岩崎、佐々木、Hossain、岡村)。
2.電位依存性プロトンチャネルの動作原理と生理機能
VSOPは電位センサードメインのみを有しポア領域をもたない(VSOP=voltage sensor-only protein)にも関わらず、電位依存性プロトンチャネル活性をもつ。VSOPはマクロファージなど免疫系の細胞に多く発現し、膜電位を介する活性酸素の産生や細胞内環境の制御に関わっていると考えられる。VSOPは、今後感染防御機構や膠原病、アレルギー、神経疾患などにおける細胞内pHや膜電位シグナルの役割を解析する上で重要なターゲット分子と考えられる。更にVSOPのシンプルな構造と機能のユニークさから、この分子動作原理の解析はイオンチャネルの動作原理一般や、様々な生命現象に関わるプロトンの透過機構一般を理解することにつながる。プロトンチャネルが膜電位を感知する機構を明らかにするため、陽電荷をもつS4を変異させ膜電位依存性の変化を検討した。3つのアミノ酸のうち細胞外側に近いアルギニンであるR1とR2が重要な働きをしていること、またアルギニンを塩基性アミノ酸であるリジンに変化させると大きな膜電位依存性のシフトが起こり、単に静電的な電荷の貢献からは説明できないなどが明らかになった。その機構はこれまで知られてきた電位依存性の電位センシングとは異なる可能性が示唆された。更に、VSOPの生物学的役割を明らかにするため、特異抗体を作成して発現様式を解析し、これまでのRNAの発現が検出されていた組織(脾臓など)でシグナルが検出された。現在細胞内局在を明らかにする実験を行っている(大河内、黒川、佐々木、高木、岡村)。
3.ゼブラフィッシュの脊髄神経回路の形成と動作原理の解明
ゼブラフィッシュを用いて、脊椎動物の脊髄神経回路の発生機構と生理機能を解析している。特定のクラスの神経細胞を、蛍光タンパク質を用いて生きたまま可視化し、それら神経細胞の発生過程を追跡することを研究手法の中心に据えて研究を続けている。平成19年は、転写因子vsx1を発現する細胞の発生に関する研究を中心に行った。その結果、vsx1は、神経前駆体細胞の最終分裂の直前で発現する遺伝子であり、また、その最終分裂で生じる2つのニューロンは常に互いに異なるクラスの神経細胞へ分化することが分かった。これは、脊椎動物中枢神経系において、異なるクラスの神経細胞が再現的に不等分裂で生じうることを示す最初の例である。
4.脊索動物型運動機能の分子基盤
無脊椎動物であるホヤのオタマジャクシ幼生は、魚類などの脊椎動物に似た遊泳 運動を行うが、その筋肉細胞が約20対しかないなど体制が著しく単純である。 カタユウレイボヤ幼生の遊泳に関わる分子機構を知るため、幼生筋で発現する nAChRを同定し、その電気特性を定めた。すると脊椎動物の筋肉型nAChRとは異な り、ホヤ幼生筋nAChRは顕著な内向き整流性と高いCaイオン透過性を示した。この分子特性が変更される変異型nAChRを作製し、これと幼生に内在するものを大幅に 入れ替えると、幼生の運動パターンは明確に変化した。この結果は、筋肉nAChR のチャネル特性がホヤ幼生遊泳に本質的な役割を持つことを示し、脊椎動物の生 理特性の成立に関わる分子進化的理解につながる機構を提示している。
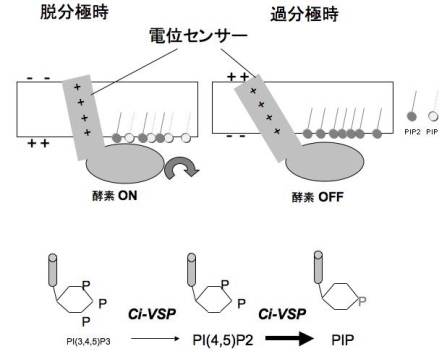
図: Ci-VSPの動作原理を図示した。脱分極時に、電位センサーの構造変化がおこり、この情報が細胞内ドメインのホスファターゼに伝わり、脱リン酸化活性(PIP3からPIP2およびPIP2 からPIPへ)がONとなり、PIP3とPIP2の濃度が減少する(上図)。VSPのホスファターゼは、おそらくこれまで知られてきたPIP3だけでなく PIP2に対しても基質として脱リン酸化を起こすと考えられる(下図)。
9.3細胞生理研究部門
痛み刺激受容・温度受容・体温調節の分子機構の解析を富永真琴が中心となって、細胞接着・細胞運動の分子機構の解析を福見知子が中心となって進めている。また、神経細胞発生に関わるCa2+動態とTRPチャネルの解析を柴崎貢志が進めている。
痛み刺激受容の分子機構の解析
痛み冷刺激やワサビの主成分等で活性化されるTRPA1チャネルの新規有効刺激をスクリーニングし、複数種類の新規刺激を見いだして解析を進めている(論文準備中)。そのうち、防腐剤として食品や化粧品に広く用いられているメチルパラベンがTRPA1チャネルの特異的刺激剤として機能すること発表した
(Br J Pharmacol 2007)。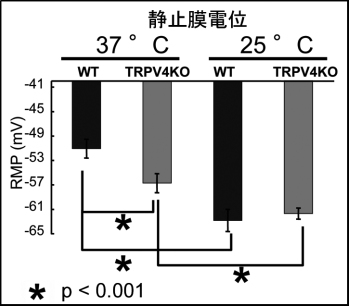
図1:メチルパラベンで用量依存的に活性化されるTRPA1電流
中枢神経における温度感受性TRPVチャネルの機能解析
海馬神経細胞のTRPV4は深部体温において恒常的に活性化して神経細胞を脱分極させ、神経の興奮性の制御に関わっていることを明らかにした (J Neurosci
2007)。行動解析・脳波解析・海馬神経細胞の長期増強解析を行っている。 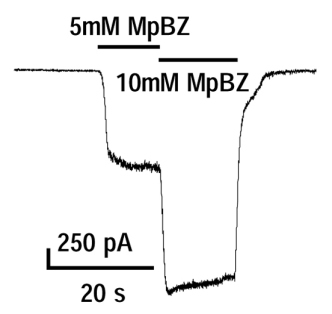
図2:25度では海馬神経細胞の静止膜電位RMPに差はないが、37度ではTRPV4欠損細胞に比べて野生型細胞で有意により脱分極している。
新規温度感受性TRPチャネルTRPM2の解析
膵臓β細胞に発現してインスリン分泌に関わっていることを明らかにしたTRPM2の機能制御物質をスクリーニングして、2-APBが強いTRPM2阻害活性を示し、膵臓からのインスリン分泌も阻害することを発見した (Br J Pharmacol, in press)。TRPM2欠損マウスの解析を進めている。
酸味受容体の構造・機能解析
PKD2L1/PKD1L3ヘテロマーが酸刺激(pH 3.0以下)に暴露されないと活性化しないが、チャネル開口は酸刺激をoffにしたときに起こる(off channel)ことを明らかにした(論文投稿中)。
ショウジョウバエPainlessの解析
ショウジョウバエで侵害性熱刺激受容に関わると推定されているTRPA1ホモログPainlessが42度以上の熱によって活性化する熱活性化チャネルであり、細胞内Ca2+濃度によって活性が制御されることを明らかにした(論文準備中)。
表皮ケラチノサイトに発現する温度受容体TRPV3, TRPV4の解析
TRPV4がケラチノサイトで細胞骨格蛋白質と結合してケラチノサイトの細胞接着ひいては皮膚のバリアー機能に関与することを明らかにした(論文投稿中)。また、ケラチノサイトで感知された温度情報が拡散性分子を介して感覚神経に伝達されると仮定して、バイオセンサーを使って温度刺激によってケラチノサイトから放出される分子を同定した(論文投稿中)。
神経発生における温度感受性TRPチャネルの役割の解析
マウス胎生期脊髄および後根神経節における温度感受性TRPチャネルの発現解析を行い、異なった時期に発現することを見いだした。また、温度受容以外の胎生期特異的な機能を有することも判明した(論文準備中)。
細胞運動の分子機構の解析
低分子 G蛋白質Rhoの標的蛋白質である mDia を介する新たな情報伝達経路の解析によって、細胞運動の時・空間的制御機構を解明することを目指している。MDiaと結合することを見いだしたDIPのノックアウトマウスの解析を進めている。