1 分子生理研究系
1.1 神経機能素子研究部門
神経機能素子研究部門では、イオンチャネル、受容体、G 蛋白質等の構造と機能に関する研究を展開している。具体的には、(1) 膜電位センサーを持たないATP受容体チャネルの、膜電位とATP濃度に依存するゲート機構の解析、(2) 代謝型グルタミン酸受容体の動的構造変化とシグナリングの多様性、および、GABAB受容体との分子会合による機能修飾の解析、(3) KCNQ K+ チャネルの機能調節機構と構造機能連関、(4) 内耳外有毛細胞のモーター蛋白プレスチンの単粒子構造解析、および分子複合体の同定、(5) 細胞膜上に発現するカフェイン受容体の分子同定と機能解析、(6) 小脳 lobule10 において特異的に見られるシナプス電流の解析等を主たる研究目標とし、学際的アプローチにより研究を進めている。2008年に発表した論文のうち代表的なもの、 Nagatomo K & Kubo Y (2008) Caffeine activates mouse TRPA1 channels but suppresses human TRPA1 channels. Proc Natl Acad Sci USA 105: 17373-17378. の内容を以下に紹介する。
我々は、マウス消化管由来のSTC-1細胞に、カフェインを投与した時に見られる [Ca2+]i の増加が、細胞外Ca2+を除くと消失することを見いだし、「カフェインによって活性化されるCa2+透過性を持つチャネル」 の存在を想定した。そして、in vitro 発現系を用いて種々の機能スクリーンを行い、マウスのTRPA1 チャネルがカフェインによって活性化されることを明らかにした。次のステップとして、この知見が神経細胞や個体においても観察されるか否かを明らかにするために、まず、TRPA1が内在的に発現しているマウスのコウコン神経節 (DRG) 神経細胞を急性単離培養し、Ca2+ i イメージングを行った。野生型マウス (WT) のDRG神経細胞では、カフェイン投与により [Ca2+]iの上昇が起きたが、TRPA1ノックアウト(KO) マウスのDRG神経細胞では、応答は観察されなかった。次に、個体レベルでも、マウスがTRPA1チャネルを介してカフェインを認識しているか否かを明らかにするために、二瓶法を用いて、水、およびカフェイン水の飲水量を測定する実験を行った。WTマウスではカフェイン水の消費量が有意に少なかったが、TRPA1-KOマウスでは違いは観察されなかった。以上の結果から、マウスの神経細胞や個体においても、TRPA1 チャネルがカフェインによって活性化されることが示された。さらに、ヒトの TRPA1 チャネルについても in vitro 発現系において機能解析を行ったところ、マウスの場合と異なり、カフェインにより、電流が抑制されることが明らかになった。
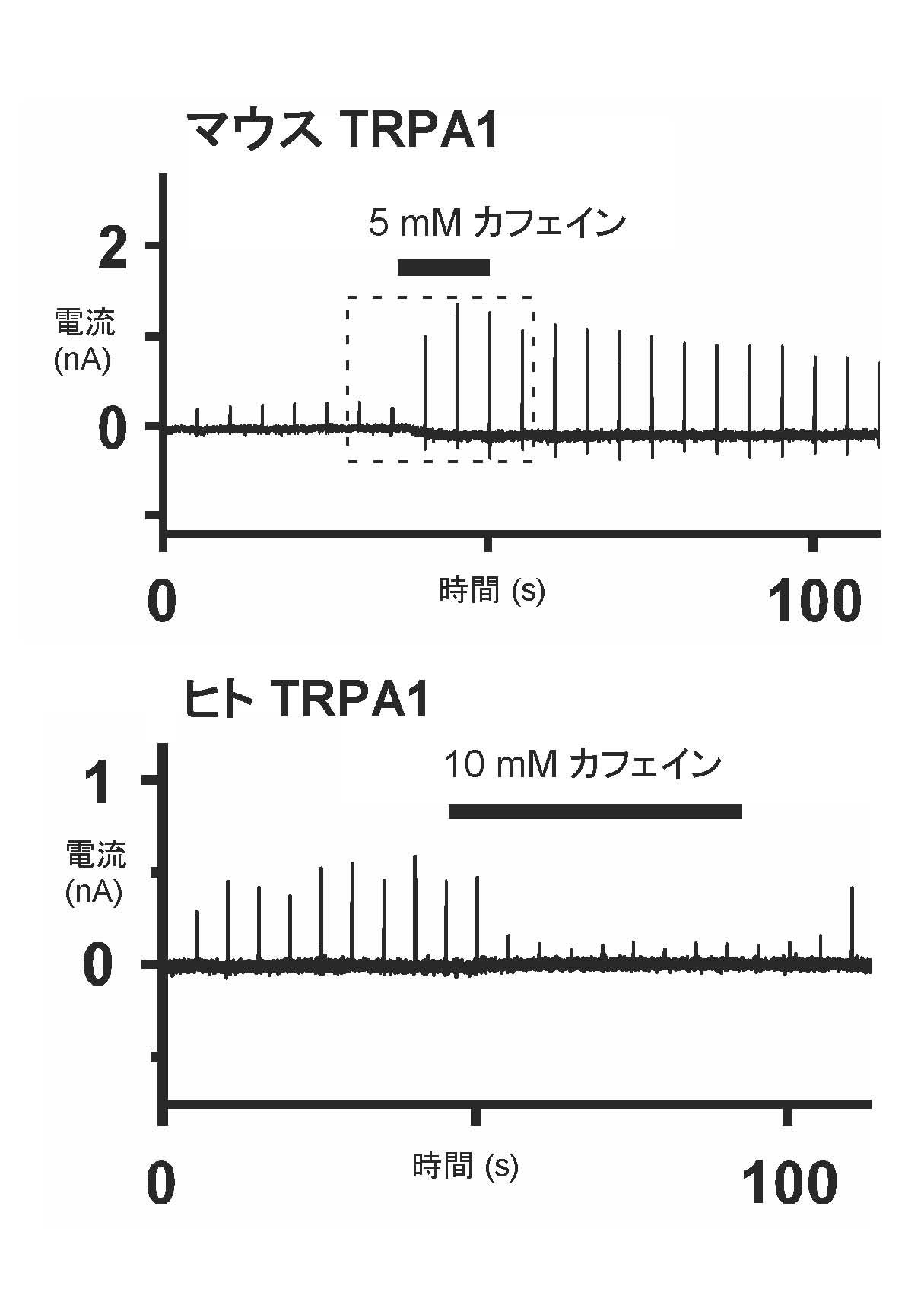
図1. 遺伝子導入したHEK293細胞のパッチクランプ法による記録。膜電位固定下で、ランプ波形の電位刺激を繰り返し与え、電流応答を記録した。 (上) マウスTRPA1 チャネルを発現させた細胞からの記録。5 mM カフェインの投与により、チャネル電流が増加した。(下)ヒトTRPA1 チャネルを発現させた細胞からの記録。10 mM カフェインの投与により、チャネル電流が減少した。
1.2 分子神経生理研究部門
哺乳類神経系の発生・分化、特に神経上皮細胞(神経幹細胞)から全く機能の異なる細胞種(神経細胞、アストロサイト、オリゴデンドロサイトなど)が分化してくる機構を研究している。このような発達における研究から、グリア細胞の多様性とその機能解析にアプローチしようとしている。近年、成人脳内にも神経幹細胞が存在し、神経細胞を再生する能力を有することが明らかとなった。この成人における神経幹細胞数の維持機構についても研究している。また、得られた新しい概念や技術は臨床研究への応用を視野に入れながら、病態の解析にも努力している。 糖蛋白質糖鎖の解析法を開発し、その生理学的意義について検討している。
神経幹細胞の発生:哺乳類脳において神経幹細胞が発生してくる分子機構の解析を行っている。早期胚から培養下で未分化神経幹細胞を単離し、神経幹細胞へと誘導する実験系を開発したので、この系を用いて未分化神経幹細胞から神経幹細胞への分化過程で働いている遺伝子を探索した。その結果、glial cells missing (gcm)遺伝子がこの段階に重要な役割を担っている可能性が示唆された。Gcm1およびgcm2のノックアウトマウスを用い、gcm1/2遺伝子の役割をさらに解析中である。
中枢神経系の細胞分化:転写調節因子Olig2の前脳形成における機能を明らかにするため、Olig2発現細胞の分化特性とOlig2欠損マウス前脳に見られる表現型を調べた。終脳では、胎生中期(~E12.5)のOlig2発現細胞はGABAニューロンに分化したが、胎生後期のものはオリゴデンドロサイトのみならずアストログリアにも分化する傾向にあった。この点に注目して胎生後期(E18.5)のOlig2欠損マウスの表現型を解析した。その結果Olig2欠損マウスでは野生型マウスと比較して、グリア型グルタミントランスポーター(GLAST)やFGF受容体3を発現するアストログリア前駆細胞の単位面積当たり数が、大脳皮質において50%程度まで減少していた。一方、前脳基底部においては、そのような変化は認められなかった。Olig2欠損マウスでは、オリゴデンドロサイトがほとんど形成されないことと合わせて、Olig2が大脳皮質ではマクログリア細胞の数と質を決める重要な因子であることが明らかにされた。
成体脳における神経幹細胞の維持機構:慢性ストレスや向精神薬投与下での神経幹細胞の維持機構を解析することにより、成体脳における神経幹細胞の存在意義を調べている。Nestin promoter/enhancer-CreERやテトラサイクリンシステムを用いたトランスジェニックマウスを作製し、神経幹細胞の動態や動物の行動変化の解析を行っている。)
脱髄機構の解析:脱髄巣において髄鞘が再生している時だけミクログリアに発現する遺伝子(シスタチンF)を見いだしたので、その機能解析をしている。
アストロサイトの機能解析:アストロサイトからグルタミン酸やATPが放出され、それらが神経活動を制御することが知られているが、その放出ダイナミクス、分子メカニズムについては全く知られていない。培養アストロサイトからのグルタミン酸放出、およびATP放出をリアルタイムで観察可能なイメージングシステムを構築した。アストロサイトからグルタミン酸、ATPが自発的に放出されること、ATPがグルタミン酸放出を引き起こすこと、グルタミン酸がATP放出を引き起こすことを明らかにした。
脳内の糖鎖解析: 糖蛋白質の微量化に努め、2次元電気泳動した後のサンプルから糖鎖を解析することを可能とした。
1.3 細胞内代謝研究部門
細胞内代謝研究部門(客員部門)では、主として機械刺激受容時や受精時における細胞内シグナリング機構、あるいは細胞のプロトンシグナリングを電気生理学とライブイメージング法を用いて調べている。また機械受容と神経機能を関連づけるシーズ研究として神経ステロイドとシナップス可塑性の研究を行っている。
細胞における機械刺激受容機構の研究
主要課題の一つは、細胞メカノセンサーの代表格である機械受容(MS)チャネルのゲーティング機構の解析である。今年度は細菌のMSチャネルであるMscLのゲーティング機構について解析した。これまでにMscLの閉構造は分かっていましたが開構造は謎であった。常時開口していると思われるミュータントはあるが、これを大腸菌に発現すると致死的であるため、大量の開状態MscL蛋白質が得られないためである。そこで我々は開チャネルミュータント蛋白質を試験管合成してその構造を電子顕微鏡で観察し、それを元にMscLゲーティングのモデル(IRISモデル)を提唱した(Yoshimura et al., PNAS,105(10):4033-4038, 2008)。このモデルを検証するために分子動力学モデルを構成し、MscLがどのようにして膜張力を受容して開口するかの全プロセスをシミュレーションしている(Sawada et al., in preparation)。このモデルは野生型や種々のミュータントの実験結果を良く説明するので、我々はMscLゲーティング機構の究極理解に近づきつつあると思っている。(曽我部)。
接着性細胞は細胞内外の力学環境(細胞が発生する収縮力や基質の硬さなど)に応じて細胞骨格や接着構造を変えて最終的に形態の変化を惹起する。しかしこのときの力学環境の感知機構は不明で、最終応答に至るシグナル機構は謎のままである。我々は、力学的負荷により、接着斑でのアクチンの重合が活性化することを発見し、このアクチン重合の活性化には、接着斑タンパク質zyxinの力学的負荷に応答しての接着斑へのリクルートが関わっていることを明らかにした(Hirata et al., J Cell Sci,121:2795-2804, 2008)。現在、力学負荷依存的にzyxinと相互作用するタンパク質の同定を進め、接着斑におけるメカノトランスダクション機構を明らかにしようとしている(平田)。
ウニ卵受精時の一酸化窒素(NO)の増加と受精膜硬化の研究
ウニ卵受精時の一酸化窒素(NO)増加について卵活性化の引き金なのか、どんな働きがあるのか検討した。NO増加と内外イオンの関係を調べるために電位固定法とNO感受性蛍光色素によるイメージングとの同時測定を行った。NO増加は受精の引き金ではなく細胞内カルシウム増加の後に増加することを確認した。受精膜硬化を抑制することが報告されているCN-やNADPH産生酵素の阻害剤DPIもNO増加を完全に抑制した。このことからNOの働きとして受精膜硬化が強く示唆されたので、この観点で研究を展開した。またNO吸収剤PTIOを用いて、NO増加を抑制した時とそうでない時とで酸素消費、Redox変化(NADH/NADPH)、H2O2 産生を測定した。酸素消費を酸素電極で測定したところ、PTIO存在下では全体の酸素消費量はコントロールに比べて減少していた。NADH/NADPHの自家蛍光シグナルもPTIO存在下ではその増加が抑制された。また酸素消費の原因であるH2O2 の産生を蛍光試薬Amplex Redで測定したところ、PTIO存在下ではH2O2 の産生は抑制されていた。さらに、PTIO存在下では化学的機械的刺激により受精膜はコントロールに比べて遥かに弱くなっていた。これらのことからNOの役割として受精膜硬化を促進する働きについて論文にまとめ発表した(Dev Biol, 322; 251-262)(毛利)。
プロトンシグナリング機構の研究
プロトンは様々な細胞機能に影響を与える重要なシグナルイオンである。破骨細胞はプロトンポンプ(V-ATPase)によって酸を分泌し骨組織を融解し(骨吸収)、骨リモデリングや生体Ca代謝の調節に貢献している。破骨細胞では骨から溶け出したカルシウム(Ca)に暴露されると骨吸収を抑制するネガティブフィードバック応答が起こり行き過ぎを防いでいるが、そのCa感受機構はよくわかっていない。私達は、これまでに高濃度のCaに暴露されるとV-ATPase活性が抑制されることを見出していたが、同時にendocytosisが促進されること、この過程がV-ATPaseのブロッカーで抑制されることが明らかにした。私達はV-ATPaseが細胞内小胞と細胞膜の間を移動して活性を調節しており、Caによって細胞膜から細胞内への移行が促進し酸分泌能が低下することが骨吸収抑制の一端を担っているのではないかと考えている。これらの結果は投稿中で、現在、V-ATPase取り込みに関わる更に詳細なメカニズムを検討している(久野)。
神経ステロイドによる脳虚血障害の保護作用
近年種々のステロイドホルモンが中枢で合成され、様々な機能を発揮することが分ってきた。特に学習記憶行動の促進やアミロイド・脳虚血による神経障害の保護作用が注目を集めているが、その分子・シナプス機構は不明である。我々は、海馬スライス標本に膜電位イメージングを適用して、代表的神経ステロイドであるエストロゲンやPREGS、DHEAなどの作用機序を調べている。本年度はDHEAの神経保護作用について解析した。閉経前の女性は閉経後の女性や同年齢の男性に比べて脳卒中の発症率が低く予後も良好であることが知られている。そこで性ホルモンが脳卒中の予防や予後に効果があると考えられ、事実そうであることが分かっている。しかし何時発症するか分からない脳卒中のために性ホルモンを投与し続けることは危険である。我々は卒中に伴う虚血による神経細胞死が発症後2-7日に亘る遅延性であることに注目し、発症後にステロイド(DHEA)を投与しても神経細胞死を防げるのではないかと考えた。結果は、予想通りで、一過的脳卒中において発症後6時間から48時間の間であれば、DHEAの一回投与で神経細胞死をほぼ完全に防止できることが分かった(Li, et al., J Cerebral Blood Flow Metabol, in press)。この手法が人間にも適用できれば大きな福音である。現在治験の可能性を検討している(曽我部)。
1.4 ナノ形態生理研究部門
1. 位相差電子顕微鏡の開発と応用(永山G)
昨年に引続き、位相板の開発と生物への応用研究を行った。位相板の開発としては、アハラノフーボーム(AB)効果を利用する無損失位相板の開発に注力した。フォーカスイオンビーム(FIB)装置を用いて極小棒磁石によるヒルベルトAB位相板が完成した(特許出願中)。 2006年10月から開始したCRESTプロジェクト「ns-nm分解能電子・光子ハイブリッド顕微鏡の開発」において、並行して3つの電子顕微鏡技術の開発を行っている(光電子銃、電子・光子ハイブリッド顕微鏡、雰囲気セル)。同一試料、同一視野に対し電顕と光顕の同時観察を行い、光顕の特性と電顕の特性をシームレスに連携させる手法を開発することが主目的である。 数年来幅広く共同研究を行い、種々の生物試料を観察してきたが、医学生物学への応用として以下のテーマに絞り込んだ。i)蛋白質の単粒子解析(Danev & Nagayama, J. Str. Biol., 2008), ii)ウィルス(Yamaguchi et al., J. Str. Biol., 2008)、バクテリアのその場観察と核酸動態(Furuhata et al. J. Nanosci. Nanotech., 2008; Nitta et al., J. Microsc., 2008), iii)人工細胞および培養細胞のその場観察と細胞内動態(Yui et al. Longmuir, 2008; Sasaki et al., Anal. Bioanal. Chem., 2008)。
i)に関しては、位相差法と従来法の比較のためGro ELを標準蛋白質として解析を行い、位相差法の優位性を示した。ii)については、重原子標識によるDNAの局在観察手法(10 nm分解能)の開発を始め、シアノバクテリアのDNA局在について新知見を得た。iii)については、電顕光顕同時観察を行うための神経細胞系の開発を開始した。また、位相差法を百万ボルト電顕に拡張する試みを韓国の基礎科学研究所との共同研究として続行した。
位相差顕微鏡は、急速凍結された光顕試料直接観察できるという特徴を持つため、光顕と電顕の相関観察を容易にする。この方向の世界的動向を概観し、合わせて将来展望を行うため、2008年11月に第39回生理研コンファレンス(39th NIPS Intl. Symposium / 7th OIB Symposium)“Frontiers of Biological Imaging -Synergy of Advanced Techniques”を組織し、150名近い参加者を得た。
2.唾液分泌機構(村上G)
傍細胞輸送調節の基盤としてタイト結合直下細胞骨格の変化を凍結割断標本の電子顕 微鏡観察した(東京歯科大学と共同研究)。灌流液に細胞内に入らない蛍光色素マーカーを入れ唾液へ移行する量から傍細胞輸送の指標とした。その結果、ムスカリン受容体刺激で傍細胞輸送が増加し、これにβアドレナリン受容体刺激を重畳するとさらに傍細胞輸送が増加した。また分泌を誘発する漢方薬のうち丹参がムスカリン受容体βアドレナリン受容体を抑制しても分泌を誘発しこのとき、傍細胞輸送を増加させた(南京医科大学と共同研究)。さらにneurokinin Aも分泌を誘発し傍細胞輸送を増加させた(日本大学松戸歯学部と共同研究)。ケンブリッジ大学(傍細胞輸送調節のシミュレーション)、カリアリ大学(HRSEMによる開口分泌の観察)、カトリック大学ローマ校(質量分析による唾液蛋白分析)等の共同研究も継続した。
3. エンドソーム-ゴルジ細胞内膜系の生理機能(大橋G)
エンドソーム-ゴルジ細胞内膜系の担う生理機能について研究を行った。細胞接着分子の発現制御や発生シグナル伝達制御におけるこれらの膜系の関与を示す新たな知見を得た。
4. 構造変化型ユニバーサル塩基の開発(片岡G)
相対する塩基が特定の塩基でなくともDNA二重鎖の形成を維持する核酸塩基、pyrimido[4,5-d]pyrimidine-2,4,5,7-(1H, 3H, 6H, 8H)-tetraoneを開発した。この人工の核酸塩基は分子内水素移動による迅速なケト-エノール互変異性化と、プリン型-ピリミジン型塩基構造の配座異性化によって、相対する塩基に呼応して構造変化し、アデニン、グアニン、シトシン、チミンのすべてと塩基対を形成する。概念的に全く新規な核酸塩基であり、様々な生化学ツールとしての利用が期待される。