1 分子生理研究系
1.1 神経機能素子研究部門
神経機能素子研究部門では、イオンチャネル、受容体、G 蛋白質等の構造と機能に関する研究を展開している。具体的には (1) ATP受容体チャネルの、膜電位とATP濃度に依存するゲート機構、(2) 代謝型グルタミン酸受容体およびGABAB受容体の動的構造変化とシグナリングの多様性、(3) KCNQ K+ チャネルの機能調節機構と構造機能連関、(4) 内耳外有毛細胞のモーター蛋白プレスチンの単粒子構造と動的構造変化、(5) マウスとヒトの TRPA1チャネルのカフェインに対する応答の相違の一次構造の基盤、(6) 小脳 lobule10 において特異的に見られるシナプス電流の分子基盤等を主たる研究目標とし学際的アプローチにより研究を進めている。2009年の発表論文のうち代表的なものKeceli B & Kubo Y (2009) Functional and structural identification of amino acid residues of the P2X2 receptor channel critical for the voltage- and [ATP]-dependent gating. J Physiol 587: 5801-5818.の内容を以下に紹介する。
我々は、これまでに、細胞外ATP によって活性化されるP2X2チャネルが、分子内に膜電位センサー領域を有しないのに、膜電位とATP に依存するゲートを示すことを明らかにした。本研究では、膜電位に依存するゲートステップの一次構造上の基盤を明らかにすることを目的として、変異体解析を行った。(1) まず、ATP結合部位と同定されている領域の変異体の解析を行ったところ、K308Rでは、電荷が保存されているにも関わらず、コンダクタンス-膜電位関係が過分極側に大きくシフトしており、また、活性化速度が速かった。この性質は、ATP結合ステップと膜電位依存的ゲートステップからなる3ステートモデルにおいて、ゲートステップのkoffを増加させることによりシミュレートできた。(2) 次に、ATPによる活性化に関与することが知られている膜貫通部位の細胞外側端に位置するアミノ酸残基の変異体の解析を行った。T339S等は、低いATP濃度では遅い膜電位依存的活性化を示し、高いATP濃度では、膜電位に依存しない恒常的活性化を示した。この性質は、3ステートモデルにおいて、ゲートステップのkoffを減少させることによりシミュレートできた。(3) koff に逆向きの変化を与えたK308RとT339Sの2重変異体の解析を行ったところ、野生型P2X2に極めて近い性質を示した。以上の実験結果から、ATP受容体チャネルP2X2の膜電位依存的ゲートには、ATP- ATP結合部位複合体と、膜貫通部位細胞外側端が複合的に寄与していることが示された。ATP- ATP結合複合体がリンカー部分を経由して膜貫通部位細胞外側端に間接的に作用し、ゲート開口につながる膜電位依存的な構造変化をトリガーすることが示唆された。
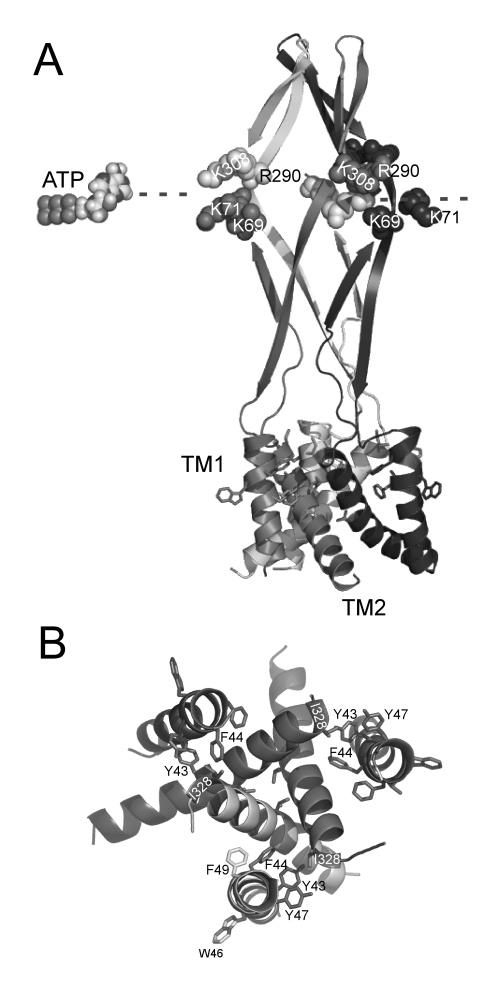
図5.1. 変異体解析により同定した膜電位依存的ゲートに重要なアミノ酸残基。Zebra fish P2X
4 の結晶構造に基づいたhomology modeling による Rat P2X2の構造上にマップした。(A) 側面図。変異体解析により同定した、ATP 結合部位近辺に位置し、膜電位依存的ゲートに重要なアミノ酸を示した。(B)
膜貫通部位を細胞外側から観察した図。膜貫通部位の細胞外側よりに位置し、膜電位依存的ゲートに重要なアミノ酸を示した。
1.2 分子神経生理研究部門
概要
哺乳類神経系の発生・分化、特に神経上皮細胞(神経幹細胞)から全く機能の異なる細胞種(神経細胞、アストロサイト、オリゴデンドロサイトなど)が分化してくる機構を研究している。このような発達における研究から、グリア細胞の多様性とその機能解析にアプローチしようとしている。近年、成人脳内にも神経幹細胞が存在し、神経細胞を再生する能力を有することが明らかとなった。この成人における神経幹細胞数の維持機構についても研究している。また、得られた新しい概念や技術は臨床研究への応用を視野に入れながら、病態の解析にも努力している。
糖蛋白質糖鎖の解析法を開発し、その生理学的意義について検討している。
1) 神経幹細胞の発生: 早期胚から培養下で未分化神経幹細胞を単離し、神経幹細胞へと誘導する系を用いて未分化神経幹細胞から神経幹細胞への分化過程でglial cells missing (gcm)遺伝子が重要な役割を担っていることを示した。今年はgcm1/2遺伝子がNotch経路と関わりを持って神経幹細胞の誘導を行っていることを明らかにした。また、今まで機能の知られていなかったFucTX蛋白質を新規フコース転移酵素として同定し、そのノックダウンが神経系の分化異常を引き起こすことを明らかにした。
2) 中枢神経系の細胞分化:転写調節因子Olig2の中脳形成における機能を明らかにするため、Olig2発現細胞の分化特性を調べ、視床網様核の形成細胞であることを示した。また、ニワトリの系を用いてグリア細胞の系譜を調べる方法をほぼ確立した。
3) 脱髄機構の解析:脱髄巣において髄鞘が再生している時だけミクログリアに発現する遺伝子(シスタチンF)を見いだしたので、その機能解析をしており、その遺伝子発現を自由に制御する系を確立した。
4) アストロサイトの機能解析:アストロサイトからグルタミン酸やATPが放出され、それらが神経活動を制御することが知られているが、その放出ダイナミクス、分子メカニズムについては全く知られていない。培養アストロサイトからのグルタミン酸放出、およびATP放出をリアルタイムで観察可能なイメージングシステムを構築した。アストロサイトからグルタミン酸、ATPが自発的に放出されること、ATPがグルタミン酸放出を引き起こすこと、グルタミン酸がATP放出を引き起こすことを明らかにした。
5) 脳内の糖鎖解析: 糖蛋白質の微量化に努め、2次元電気泳動した後のサンプルから糖鎖を解析することを可能としたので、髄鞘蛋白質MOGの糖鎖構造を決定した。さらに、末梢神経系における髄鞘糖鎖の解析を行った。
1.3 ナノ形態生理研究部門
1. 位相差電子顕微鏡の開発と医学生物学への応用(永山G)
機構内連携研究、一般共同研究、計画研究及びCRESTにおいて位相差電顕の開発と応用を行っている。機構内連携では位相差トモグラフィーを軸とした分厚い生物試料の立体構造解析法開発と脳神経解剖学への応用を、CRESTでは並行して3つの電子顕微鏡技術の開発を行った(光電子銃、電子・光子ハイブリッド顕微鏡、雰囲気セル)。同一試料、同一視野に対し電顕と光顕の同時観察を行うハイブリッド顕微鏡および雰囲気セルの開発は順調に進んだが光電子銃開発は壁に当たっている。
共同研究では位相差電顕の医学生物学への応用として、ⅰ) 蛋白質の単粒子解析、ⅱ) ウィルス、バクテリアのその場観察と核酸動態、ⅲ) 人工細胞および培養細胞のその場観察と細胞内動態、ⅳ) 脳組織の凍結切片の無染色観察を行った (J Str Biol)。ⅰ) に関しては、位相差法と従来法の比較のためGro ELを標準蛋白質として解析を行い、位相差法の優位性を示した (Ultramicroscopy & J Biol Chem)。ⅱ) については、重原子標識によるDNAの局在観察手法開発を行い(J Membr Biol)、シアノバクテリアのDNA局在について新知見を得た (J Microscopy)。ⅲ) については、電顕光顕同時観察により人工細胞内のDNA局在を明らかにした (Biomaterials)。
2.(村上G)
ⅰ) 傍細胞輸送の形態学的生理学的基盤:水分を大量に分泌する唾液腺では60%以上の水分がタイト結合を越えて分泌される(傍細胞輸送)。無化学固定試料の凍結割断試料を電子顕微鏡観察によりclaudin索と細胞骨格の微細構造を明らかにし、傍細胞経路の分子フィルタサイズを計測した。2008年には傍細胞開閉のCa依存性を示した。2009年には傍細胞輸送の駆動力に静水圧の寄与を検討した。そのため灌流動脈圧を測定しながら、灌流流速を変化させ、水分泌速度、蛍光マーカー(Lucifer Yellow)の分泌を測定した。その結果、灌流圧に応じて、水分泌も蛍光マーカー分泌も変化し、傍細胞輸送の駆動力に静水圧が寄与することが判明した。
ⅱ) 漢方薬の唾液水分分泌増強作用機構:唾液腺の水分分泌を増加させる漢方薬のうち、この漢方薬のみで傍細胞経路の開閉を起す丹参(DS)について、小さい分子を通す傍細胞経路がより活性化されることが見つかった。水分泌反応の用量依存性と反応の潜時を調べ、受容体が唾液腺細胞の表面ではないことが示唆された。DS水溶液のHPLCパターンを検討し経口投与濃度と血液濃度の関係を検討した。
3. エンドソーム-ゴルジ細胞内膜系の生理機能(大橋G)
エンドソーム・ゴルジ細胞内膜系の担う生理機能について研究を行った。 平面細胞極性シグナル伝達の制御において、これらの細胞内膜コンパートメント上で機能する分子を探索し、その候補分子を挙げることに成功した。
4. 構造変化型ユニバーサル塩基の開発(片岡G)
相対する塩基が特定の塩基でなくてもDNA二重鎖の形成を維持する核酸塩基、pyrimido[4,5-d] pyrimidine-2,4,5,7-(1H,3H,6H,8H)-tetraone を開発した。この人工の核酸塩基は分子内水素移動による迅速なケト-エノール互変異性化と、プリン型-ピリミジン型塩基構造の配座異性化によって、相対する塩基に呼応して構造変化し、アデニン、グアニン、シトシン、チミンのすべてと塩基対を形成する。概念的に全く新規な核酸塩基であり、様々な生化学ツールとしての利用が期待される。