4 統合生理研究系
4.1 感覚運動調節研究部門
高次脳機能(顔認知など)に関連する脳反応、各種感覚や運動に関連する脳反応などを、各種ニューロイメージング手法(脳波、脳磁図、機能的MRI、近赤外線分光法、経頭蓋磁気刺激)を用いて研究している。2010年に発表した論文のうち代表的な2研究を紹介する。
(1) Nakato E, Otsuka Y, Kanazawa S,Yamaguchi MK, Kakigi R (2010) Distinct differences in the pattern of hemodynamic response to happy and angry facial expressions in infants --A near-infrared spectroscopic study--. Neuroimage (in press).
他者の表情を読むことは、日常生活において対人コミュニケーションを円滑に行うためにとても重要である。今回は、乳児が日常接することの多い表情の中で、“笑顔”と“怒った顔”の脳反応をNIRSを用いて調べた。その結果、“笑顔”では、顔刺激の提示終了後でも脳反応の増加が継続していたのに対し、“怒った顔”では急速に脳反応が低下した(図1)。また、“笑顔”に対しては左側頭部、“怒った顔”では右側頭部で脳反応の増加が認められた(図2)。つまり、“笑顔”(ポジティブ表情)と“怒った顔”(ネガティブ表情)を、左右の別々の半球で処理していることが示された。“笑顔”は、他者に喜びの情報を伝えるため、脳の活動が継続して活動するが、一方で、 “怒った顔”は、警告や危険を示す情報を伝え次に行動を移す必要があるため、脳の活動が急速に低下していくと考えられる。つまり、生後間もない乳児が、ポジティブ表情とネガティ表情から読み取れる生物学的な意味を解釈し、その情報に応じて別々に処理している可能性が判明した。今回の研究は、赤ちゃんの脳内でポジティブ表情とネガティブ表情に反応する神経基盤を明らかにした世界で初めての研究である。なお本研究は、中央大学文学部との共同研究であり、毎日新聞、中日新聞などで研究内容が紹介された。
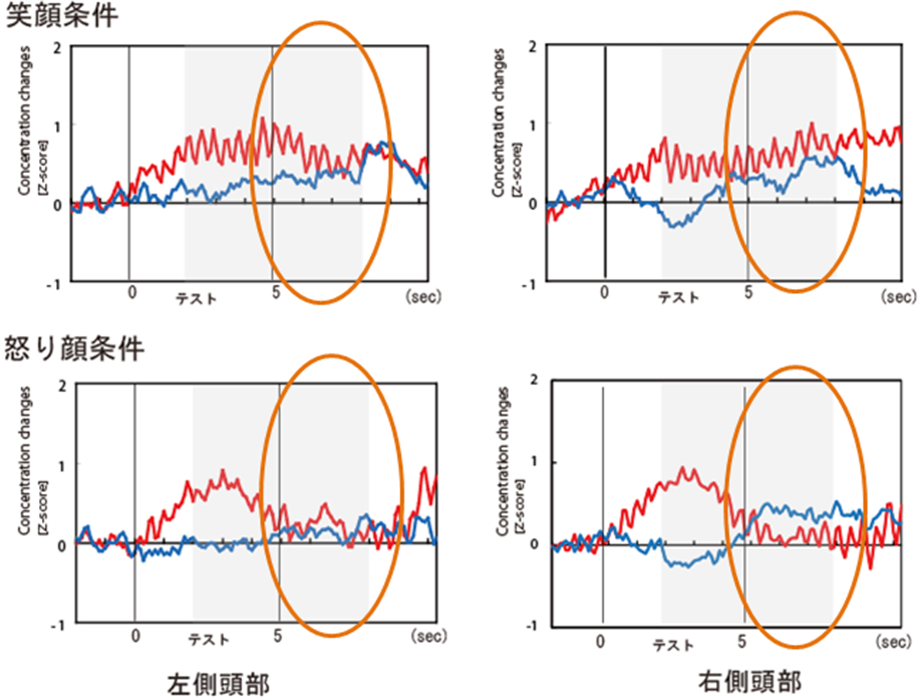
図1. 笑顔と怒った顔に対する時系列データ
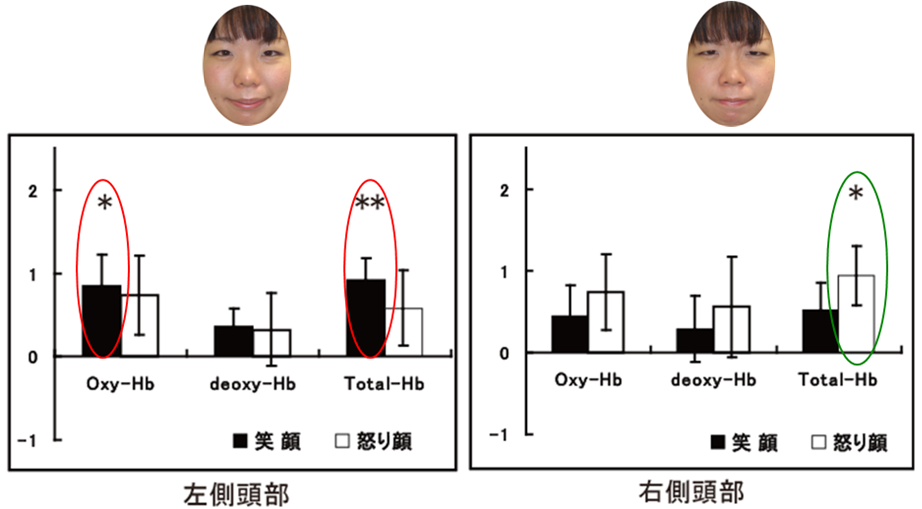
図2. 笑顔と怒った顔に対する左右側頭部での活動
(2) Inui K, Urakawa T, Otsuru N, Takeshima Y, Kakigi R
1.Non-linear lows of echoic memory and auditory change detection in humans (BMC Neuroscience 11:80, 2010).
2.Echoic memory of a single pure tone indexed by change-related brain activity (BMC Neuroscience 11:135, 2010)
感覚記憶と変化関連脳活動
脳波及び脳磁図を用いて聴覚感覚記憶の形成と減衰を検討した。図1に示すように音の変化は上側頭回に100ミリ秒付近頂点の特異的な変化関連活動を惹起する。変化の検出には現在と過去の状態の比較が必要であり、短期記憶が関与している。従ってこの変化関連活動を指標にすることで間接的に記憶状態を観察することができる。音変化の前に呈示する基準音の長さを25ミリ秒から1000ミリ秒まで変化させたところ、変化関連活動の振幅は基準音の長さの対数に比例して増加した。基準音と変化音の間のブランクを1ミリ秒から1000ミリ秒まで変化させた場合、変化関連活動の振幅はブランクの長さに対数に比例して減少した。このことから、聴覚感覚記憶の形成と減衰はいずれも時間の対数で表現されることが示された。このような感覚記憶に基づく変化検出システムは、変化に対して迅速に対応するためのリアルタイムモニターであると考えられる。従来変化関連活動(ミスマッチ反応)は、繰り返し呈示される基準音の中に時折逸脱音を混入させることで記録されてきた。しかしながらこのような手法はリアルタイムモニターの概念とマッチしない。そこで次に、単一の純音呈示が感覚記憶を形成し、その後に続く逸脱音が変化関連活動を惹起するかどうかを検討した。結果は非常に明瞭な上側頭回の変化関連活動を示した。上側頭回は「変化」発生時に開くリアルタイムsensory gate であると考えられる。
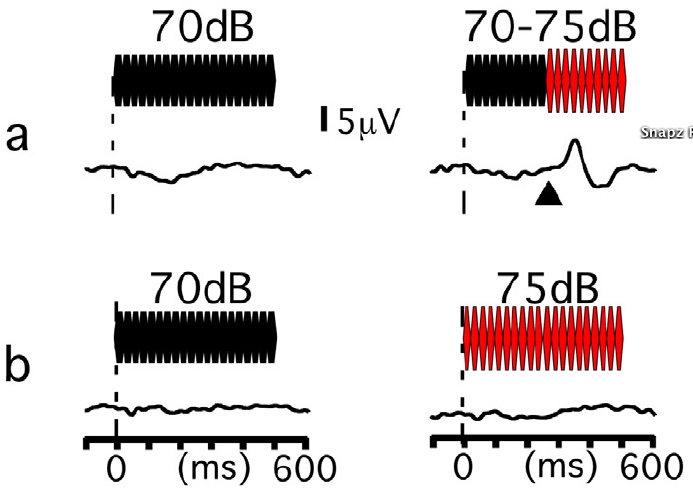
図3. 聴覚変化関連活動。a) 600msの基準音に対する脳反応(左)と、300msの基準音(70dB)の後に5dB大きい音が続く場合の脳反応(右)。三角は変化発生のタイミング。変化発生から約100ミリ秒後頂点の明瞭な変化関連活動が惹起される。一方70dBと75dBの音を交互に呈示した場合(b)には変化活動はなく、この活動が突然の5dBの変化によるものであることがわかる。
4.2 生体システム研究部門
本研究部門は、脳をシステムとして捉え、大脳皮質・大脳基底核・小脳・脳幹などの脳領域がいかに協調して働くことによって随意運動を可能にしているのか、そのメカニズムや、これらの脳領域が障害された際に、どのような機構によって症状が発現するのかなどの病態生理を明らかにし、さらにはこのような運動障害の治療法を開発することを目指して、霊長類やげっ歯類を用い神経生理学的手法、あるいは神経生理学的手法と神経解剖学的手法を組み合わせて研究を行っている。
2010年に発表した論文を紹介する。
Hashimoto M, Takahara D, Hirata Y, Inoue KI, Miyachi S, Nambu A, Tanji J, Takada M, Hoshi E (2010) Motor and non-motor projections from the cerebellum to rostrocaudally distinct sectors of the dorsal premotor cortex in macaques. Eur J Neurosci 31: 1402-1413.
Saga Y, Hirata Y, Takahara D, Inoue K, Miyachi S, Nambu A, Tanji J, Takada M, Hoshi E (2011) Origins of multisynaptic projections from the basal ganglia to rostrocaudally distinct sectors of the dorsal premotor area in macaques. Eur J Neurosci 33: 285-297.
小脳と大脳基底核は視床(thalamus)を介して大脳皮質に投射し、その活動を支えている。したがって、大脳皮質の特定の領域が、どのような小脳の領域あるいは大脳基底核の領域から入力を受けているのかは、重要な問題である。しかし、このような経路は多くのシナプスを介しているので、シナプスを超えない従来の標識物質を用いる方法で調べるには限界があった。狂犬病ウイルスが逆行性にシナプスを超えて感染する性質を利用して、標識物質として用いる方法が近年、注目を集めている。本論文は、マカク属サルを用い、運動の企図と実行に関係していると考えられる運動前野背側部の尾側領域(F2領域)に狂犬病ウイルスを注入し、経時的に逆行性に感染領域を調べることにより、F2に投射している小脳領域、大脳基底核領域を調べようとしたものである。さらにF2領域は、吻側部(F2r)と尾側部(F2c)では機能が異なり、F2rは運動を指示する手がかり刺激に、F2cは左右どちらの手を使うかなど運動実行に近い情報を担っていることが解ってきているので、狂犬病ウイルスをF2rとF2cに打ち分けた。
F2r、F2cに狂犬病ウイルスを注入して3日後では、主に注入側とは反対側の小脳核のニューロンに標識が見られた。F2r注入では、歯状核の尾腹側部に標識ニューロンが見つかったのに対し、F2c注入後では歯状核の吻背側部に加え中位核、室頂核にも標識ニューロンが見つかった。注入後4日目では、小脳皮質外側部のプルキンエ細胞も標識されるようになった。F2r注入では小脳半球Crus I, IIが、F2c注入ではCrus I, IIの他にCrus III, VIIIが標識された。これらの結果から、F2cはF2rに比べて広い小脳領域から入力を受けていること、F2cとF2rに出力を送っている小脳皮質と小脳核の領域は異なることが明らかになった。
一方、大脳基底核においては、F2r、F2c注入後、1つシナプスを介した2次ニューロンが淡蒼球内節(GPi)と黒質網様部(SNr)に分布していた。F2c注入では尾腹側のGPiニューロンが標識されたのに対し、F2r注入では吻尾側中央部で背側部のニューロンが標識された。SNrでは、F2c、F2r注入後、それぞれSNrの尾側と吻側に標識されたニューロンが見つかった。3次ニューロンは、淡蒼球外節(GPe)、視床下核(STN)、線条体(striatum)などに分布していた。F2c注入ではGPeの広い領域のニューロンが標識されたのに対し、F2r注入では、GPeの吻側と背側の限られた領域のニューロンが標識された。STNでもF2c注入では広い領域のニューロンが標識されたのに対し、F2r注入では腹側の限られた領域のニューロンが標識された。線条体では、F2r、F2c注入後、線条体ブリッジ領域とその周辺、および腹側線条体を含む広い領域に標識が見られた。これらの結果は、F2cとF2rに至る多シナプス性入力は、大脳基底核の出力部では分離しているが、入力部やその途中では、入り混じっていることを示しており、大脳基底核の入力部など初期の段階では、情報の収束・統合が行われていると考えられる(図参照)。
本研究は、京都大学霊長類研究所、玉川大学脳科学研究所との共同研究で行われた。
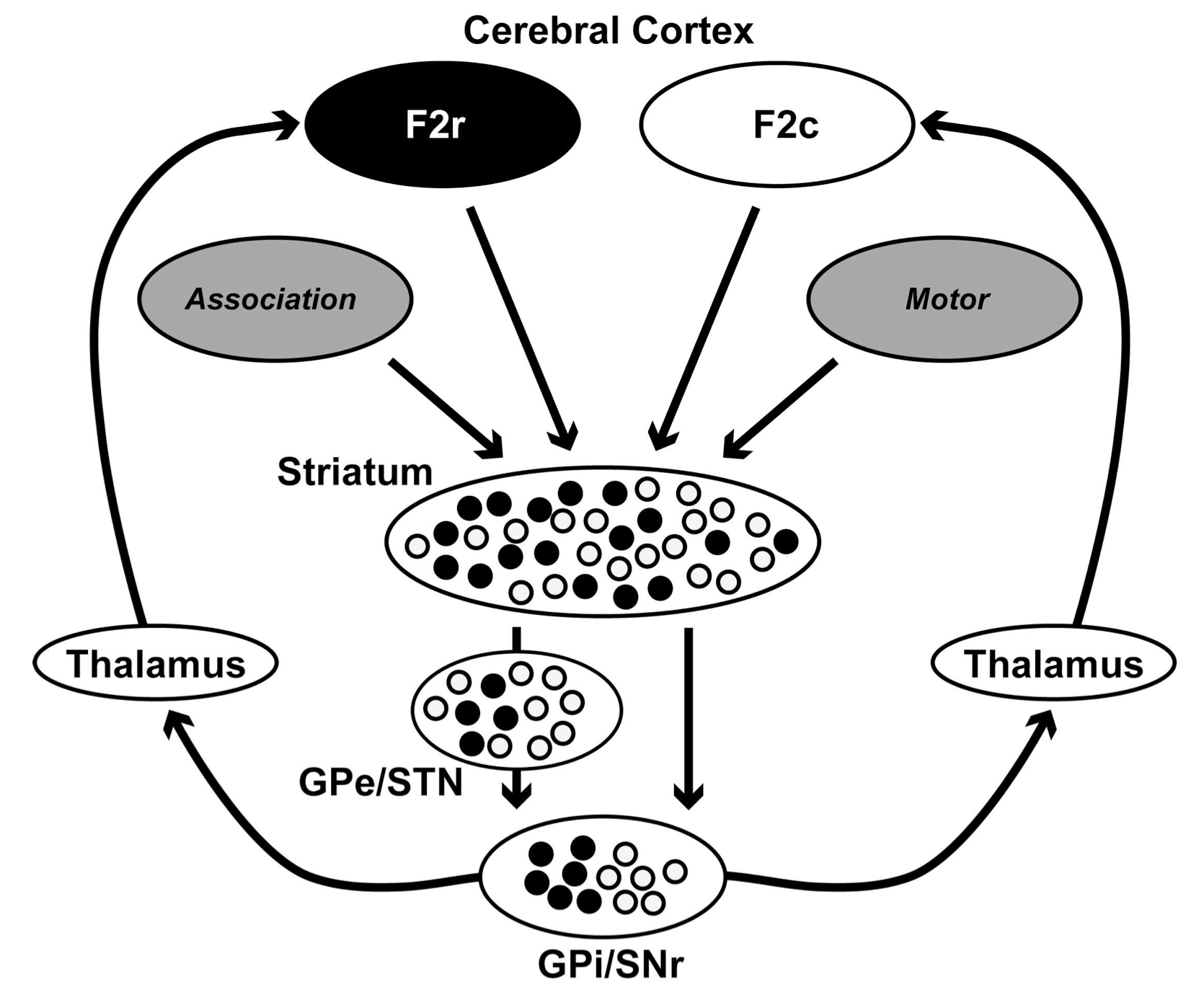
図 大脳基底核を巡る情報処理を示す模式図。
異なる大脳皮質からの情報は、大脳基底核の入力部や中継核では入り混じっている(情報の収束・統合が行われている)が、次第に出力部に近づくにしたがって分離し、それぞれ運動前野のF2r領域、F2c領域に至る。
Association, 前頭連合野;F2c, F2r, 運動前野背側部尾側領域のうち尾側部と吻側部;GPe, GPi, 淡蒼球外節、内節;Motor, 運動野;SNr, 黒質網様部;STN, 視床下核;Striatum, 線条体;Thalamus, 視床
Copyright (C) 2010 NIPS