3 生体情報研究系
3.1 感覚認知情報研究部門
感覚認知情報部門は視知覚および視覚認知の神経機構を研究対象としている。主に無麻酔のサルの視覚野に微小電極を刺入してニューロン活動を記録し、ニューロンの刺激選択性や、異なる種類の刺激への反応の分布を調べることにより、視覚情報の脳内表現を明らかにすることを試みると共に、さまざまな行動課題時のニューロン活動を分析することにより、それらの視覚情報が知覚や行動にどのように関係しているかを調べている。具体的な課題として(1)初期視覚野における輪郭とその折れ曲がりの表現、(2)下側頭皮質における色選択性ニューロンが色知覚や色弁別にどのように関わっているか、(3)高次視覚野における色情報処理経路の同定、(4)視覚関連領野における要素的な刺激のグルーピングのメカニズムに関する研究、(5)質感に関わる視覚情報の脳内表現の研究などを行った。またサルでfMRIを用いた実験により、(6)色に選択的に表現する脳領域の同定を進めその成果を論文として発表した。これは生理研のMRI装置を用いた初めてのサルのfMRI研究の発表である。2009年に上記の(3)について発表した論文を紹介する。
Yasuda M, Banno T and Komatsu H. Color selectivity of neurons in the posterior inferior temporal cortex of the macaque monkey. Cerebral Cortex, doi: 10.1093/cercor/bhp227, 2009
サルの下側頭皮質は大脳皮質の腹側に存在する高次領野で、損傷されると色の識別が障害されることからヒトで色知覚に重要な役割を果たす腹側高次視覚領野と対応するものと考えられる。我々は下側頭皮質後部(PIT)でニューロンの刺激選択特性を詳細にマッピングした結果、鋭い色選択性を持つ細胞が密集して存在し、視野の場所を表現する地図を持つ領域が存在することを発見しPITC(下側頭皮質後部色領域)と名づけた。この領域は色情報処理に深く関係しているものと考えられる。
図1Aは今回マッピングを行ったPITの場所を示す。この場所は下側頭皮質の入口にあたる場所である。図1Bは脳の写真の上に我々が新しく発見したPITCの位置を示している。PITCは後中側頭溝(PMTS)をまたがって存在し、上部のニューロンは中心視野に受容野を持つが、下部に移動すると受容野は周辺視野を含むようになり、更に後部では上視野、前部では下視野に受容野を持つという、全体として大ざっぱな視野の地図を持っていた。大脳視覚野にはいくつもの視野の地図が存在するが、別々の視野地図は別々の機能に対応すると考えられている。従ってPITCも特定の機能に関係した一つの領野に対応するものと考えられる。我々はCIE-xy色度図で一定間隔に分布した同じ明るさ(輝度)の色刺激のセットを使って、ニューロンの反応を調べた。その結果PITC内から記録されたニューロンの多くは鋭い色選択性を持っていたが、PITCの外で記録されたニューロンは鋭い選択性を示さなかった。鋭い色選択性を持つニューロンが多いことから、PITCは色情報処理に深く関係した領野であると推測される。
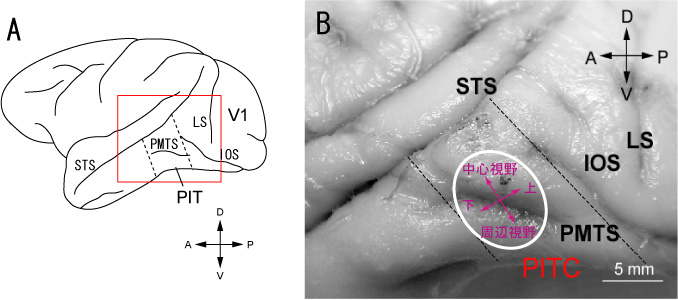
図1:A, サルの脳の外側面とPITの場所。B, PIT付近の脳の拡大写真とPITCの場所および視野表現。
図2はPITCとその周りの領域のニューロンの性質をマッピングした結果を示している。図2Aは色選択性の鋭さを示している。SとBはそれぞれ一定の定量的な基準を上回る鋭い色選択性を示した細胞(S)と示さなかった細胞(B)を表す。点線より下の領域に鋭い色選択性を持つニューロンが密集して存在していた。この部分をPITCと名づけた。図2Bは受容野が視野のどこに位置していたかを示している。PITCの上の方には中心視野に受容野を持つ細胞(F)が多いのに対し、下の方ではより周辺視野を含む受容野を持ち、更に上視野に受容野を持つ細胞(○)が後に存在し、下視野に受容野を持つ細胞(●)が前の方に存在し、全体として大ざっぱな視野地図を持っていることが分かった。この領域のニューロン活動が色知覚の成立とどのように関わっているのかを知ることが今後の課題である。
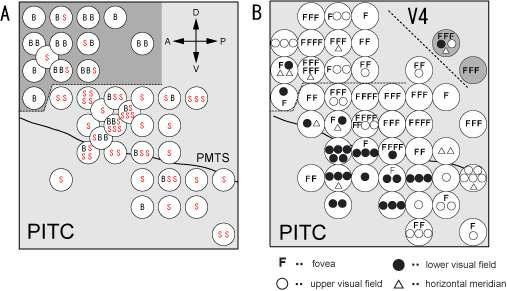
図2:A, PITC付近から記録したニューロンの色選択性の分布。Sは鋭い色選択性を示した細胞。Bは広い色選択性を示した細胞(sparseness index0.3で区分)。B, PITC付近から記録したニューロンの受容野の視野位置の分布。
3.2 神経シグナル研究部門
神経シグナル研究部門では、脳神経系の機能的素子であるイオンチャネル、トランスポーター、リン酸化酵素などの知見を基礎として、より複雑な系である神経回路の機能を理解することを目指して研究を行っている。特に2009年に古江准教授が着任後は特にin vivoの実験に重点を置き、機能分子の生体内での活動に注目して研究を行っている。また遺伝子改変マウスの行動実験も積極的に取り入れ、海馬依存性もしくは扁桃体依存性の記憶形成にリン酸化酵素がどのように関わっているかを検討している。2010年に発表された論文2編を紹介する。
Uta D, et al. TRPA1-expressing primary afferents synapse with a morphologically identified subclass of substantia gelatinosa neurons in the adult rat spinal cord. Eur J Neurosci 31:1960-1973, 2010.
TRPA1チャネルは冷覚と炎症性侵害性シグナルを伝える分子であるとされている。TRPA1は小型の急進性神経細胞の末梢神経終末と中枢側の神経終末の両方に発現している。脊髄の膠様質Substantia gelatinosa (SG)は侵害性信号の入力部位であるが、SGの神経細胞がどのような入力を受けるかは十分調べられていない。本研究では、ラット脊髄のスライス標本を用いて、TRPA1チャネルの活性化がSG神経細胞の入力にどのような影響を与えるかをパッチクランプ法を用いて検討した。TRPA1アゴニストであるcinnamaldehyde により連続的興奮性シナプス電流(EPSC barrage)が誘発された。cinnamaldehydeは抑制性電流には影響を与えず、シナプス後細胞にも影響を与えなかった。テトロドトキシン存在下でcinnamaldehydeは微小EPSCの頻度を増加させたが振幅は変化しなかった。またcinnamaldehydeはC線維の誘発EPCSを抑制した。これらの結果より、脊髄のTRPA1は一次求心性線維から垂直性細胞・放射性細胞への微小興奮性シナプス伝達を促進していることが明らかとなった。また中枢性のTRPA1が、SGの神経細胞に2方向性の調節をしている可能性を示す。
Satake S et al. Glutamate transporter EAAT4 in Purkinje cells controls intersynaptic diffusion of climbing fiber transmitter mediating inhibition of GABA release from interneurons. Eur J Neurosci 32:1843-1853, 2010.
神経伝達物質は放出後シナプス間隙から拡散して近隣のシナプスに作用し、同じ神経細胞への他の入力をコントロールする。小脳登上線維から放出される興奮性神経伝達物質(おそらくグルタミン酸)は、バスケット細胞—プルキンエ細胞シナプスでGABAの放出を抑制することが示されている。本研究では、アミノ酸トランスポータが登上線維伝達物質による抑制(CF inhibition)を調節するかを検討した。
プルキンエ細胞のトランスポータであるEAAT4を薬理学的にブロックすると、CF inhibitionは顕著に増強し、一方、登上線維のテタヌス刺激によってグルタミン酸トランスポーターを長期増強させるとCF inhibitionは減弱した。電気生理学的測定をおこなったスライス標本で免疫染色を行ったところ、EAAT4の発現レベルとCF inhibitionの間に逆比例の関係があった。また小脳小葉の間でもCF inhibitionに違いが見られ、EAAT4の発現が少ないlobule IIIではCF inhibitionが著明であり、EAAT4の発現が多いlobule XではCF inhibitionはほとんど見られなかった。これらの結果より、神経細胞に発現するグルタミン酸トランスポータは、登上線維伝達物質の拡散を調節することによりプルキンエ細胞への抑制性入力を調節していることが明らかになった。
3.3 神経分化研究部門
吉村を中心とする研究グループでは、大脳皮質視覚野の神経回路特性とその経験依存的発達メカニズムの解析を行っている。本年度はin vivo 視覚生理実験を行うシステムを立ち上げ、大脳皮質視覚野ニューロンの視覚刺激に対する反応特性の解析を開始した。また、in vitro脳切片標本にホールセルパッチクランプ法と光刺激法を適用した神経回路解析の引き続き実施している。その中で最も進展があった研究内容を以下に記す。
これまでに我々は、大脳皮質視覚野において興奮性結合がある2/3層錐体細胞ペアは、その周辺の興奮性細胞からの入力を高い頻度で共有することを見出し、非常に微細なスケールの神経回路網が視覚野内に存在することを報告した。この微小神経回路網の成熟には活動依存的メカニズムが関与するかを調べる目的で、発達期に両眼の眼瞼を縫合することにより形態視を遮断して飼育したラットの視覚野神経回路を解析した。両眼遮蔽した視覚野の2/3層錐体細胞ペアに興奮性神経結合がみられるかを調べたところ、正常な視覚体験を経たコントロール群とほぼ同様な確率で結合が検出され、結合強度にも有意な相違は見られなかった。しかしながら、この神経結合の有無にかかわらず、2/3層錐体細胞ペアが周辺の興奮性ニューロンからの入力を共有することは稀であった。従って、個々のニューロン間の神経結合は両眼遮蔽した視覚野においても形成されるが、これらは微小神経回路網を構築するようには統合されていないと考えられ、微小神経回路網の形成には生後発達期の正常な視覚体験が重要であることが示唆された。両眼遮蔽した視覚野では、視覚刺激に対する反応選択性が低下することが報告されているので、この神経回路は視覚野ニューロンの反応選択性形成に関与する可能性が考えられる。
東島を中心とするグループは、体制が比較的単純な脊椎動物であるゼブラフィッシュを用いて、脊髄神経回路の発生機構および回路機能の解析を行っている。胚期、幼生期初期には、ゼブラフィッシュの体はほぼ透明である。この利点を生かし、蛍光タンパク質を特定のクラスの神経細胞に発現させ、それら神経細胞を生きたまま可視化することを研究手法の中心に据えて研究を進めている。また、近年開発された光遺伝学ツール(チャネルロドプシンやハロロドプシンなど)も活用して神経回路の作動機構を調べている。
2010年度は転写因子Chx10を発現する細胞の解析を中心に研究を行った。これまでの電気生理学的な解析により、脊髄Chx10細胞は遊泳行動時に同側の運動ニューロンにフェージックな興奮性入力をすることを明らかにしている。Chx10細胞群の遊泳行動における役割をさらに解析するために、Chx10発現細胞にチャネルロドプシンを発現する魚を作製し、様々な領域に光照射を行った。その結果、後脳の後方部から脊髄の前方部にわたる領域において、光刺激により遊泳行動の誘発が可能であることが示された。特に後脳後半部は光刺激に最も強く反応した。また、カルシウムイメージングや電気生理学的解析により、後脳Chx10細胞が仮想遊泳行動時に活動することを明らかにした。この結果は、脊髄だけでなく、後脳後半部のChx10細胞群も遊泳行動に重要な役割を果たしていることを示唆している。
Copyright (C) 2010 NIPS