私たちの色知覚は色刺激の色(色度)だけでなく色刺激がおかれた背景の明るさに大きく影響されます。例えば同じ色でも背景が明るければ茶色に感じ、暗ければ橙色に感じるといったことが起きたり、無彩色(白、黒、灰)の色刺激では、背景より明るいときには白に見える刺激が、背景より暗いときには黒に見えるといった大きな変化が起きたりします。色の情報は目から大脳皮質一次視覚野に伝えられた後、腹側視覚経路とよばれる経路を通って処理されます。サルの大脳視覚皮質でこの経路に属するV4野や下側頭皮質(IT)は色の知覚に深く関与していると考えられており、特定の色を見ているときに強い反応を示す「色選択性細胞」が多く存在していることが知られています(図1A)。また下側頭皮質には色選択細胞がたくさん固まっている場所が前の方と後の方に存在することが見いだされており、それぞれ下側頭皮質前部色領域(AITC)と後部色領域(PITC)と名付けられています(図1B)。明るさの影響を受けて変化する色の見えのメカニズムを理解するためには、これらの場所に存在する色選択性細胞が表現する色の情報が、背景との明るさの違いによってどのように変化しているのかを明らかにすることが重要です。そのため本研究ではサルのV4野、PITC、AITCのそれぞれに存在する色選択性細胞の活動を記録し、個々の色選択性細胞の応答や色選択性細胞集団が表現する色の情報が、色刺激の輝度コントラスト(背景との明るさの違い)によってどのように変化するかを調べました。その結果、PITCの神経活動がもっとも色の見えに相関した変化を示すことが分かりました。その一方で、AITCの神経活動は明るさ情報とは切り離された色情報のみを表現していることも明らかになりました。
説明
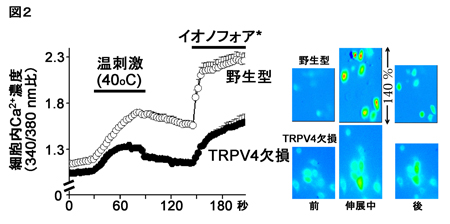
注視課題を遂行中のマカクザルに15色の色刺激をCRTモニタの背景(灰色:10 cd/m2)より高い輝度 (20 cd/m2:明るい色刺激セット)と低い輝度(5 cd/m2:暗い色刺激セット)で呈示し(図2)、微小電極を用いてサルのV4野およびPITC、AITCから色選択性細胞の活動を記録した。そして、明るい色刺激に対する神経活動と暗い色刺激に対する神経活動の比較を行った。記録された色選択性細胞の応答の例を図3に示す。このニューロンは明るい色刺激セットでも暗い色刺激セットでも赤紫色に強く応答したが、最大応答を示した色は両者の間で少しずれていた。
図1
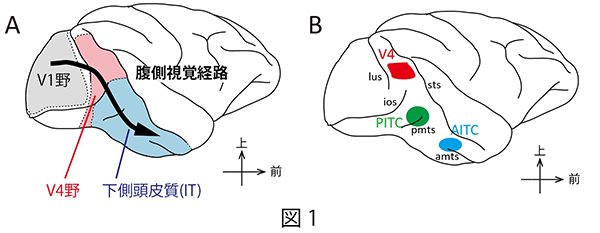
図1の説明 |
図2
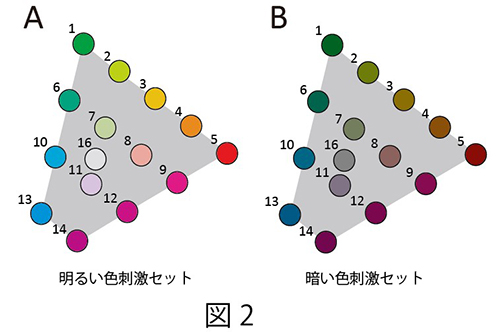
| 図2の説明 実験で用いた30個の色刺激。明るい色刺激セット(A)と暗い色刺激セット(B)。数字は同一の色度を持つ刺激対を表している。 |
図3
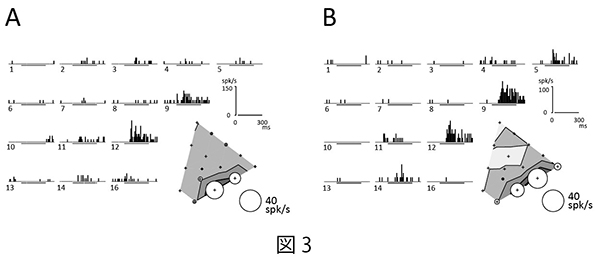
| 図3の説明 V4野から記録した赤紫によく反応した一つの色選択性細胞の応答例。神経活動をperi-stimulus-time-histogram(PSTH)(パネル左上)と円の大きさ(右下)でそれぞれ表している。A.明るい色刺激セットでは12番の色に最も強く応答した。B.暗い色刺激セットでは9番の色に最も強く応答した。AとBでは色選択性の全体的な傾向は似ているが、神経応答のピークがわずかに異なっている。数字は同一の色度を持つ刺激対を表している。PSTHは10ミリ秒のビン毎の発火頻度を示したもの。PSTH下の横線が刺激呈示期間を表す。 |
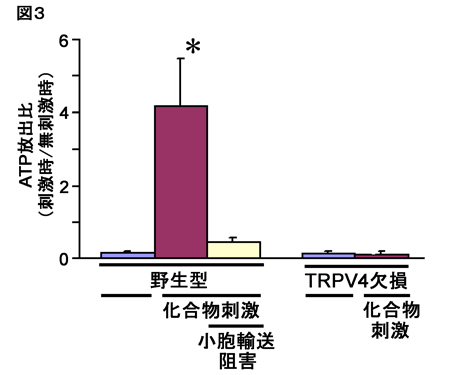
色刺激の輝度コントラストの変化が細胞の色選択的な応答に与える影響を調べるために、同一の色度の刺激対への応答を比較することで、明るい色刺激セットに対する応答強度と暗い色刺激セットに対する応答強度の相関を細胞ごとに計算した。その結果、V4野とPITCでは相関の低い細胞の割合が大きいのに対し、AITCでは大部分の細胞は高い相関を示すことが分かった(図4)。このことは、AITCでは色選択性細胞の応答に対する輝度コントラストの影響がV4やPITCに比べて小さく、輝度コントラストによらず色選択性が安定していることを示している。
図4
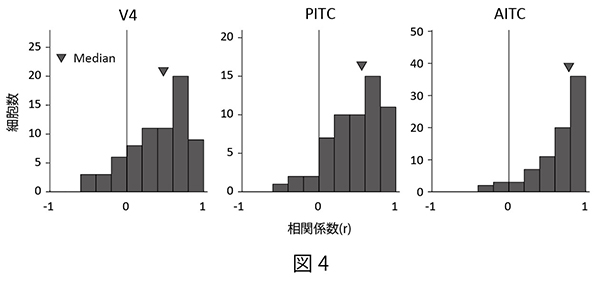
| 図4の説明 個々の色選択性細胞において、明るい色刺激セットによって引き起こされた応答強度と暗い色刺激セットによって引き起こされた応答強度から計算した相関係数の分布。相関が低いほど、色選択的応答に対する輝度コントラストの影響が大きいことを表している。 |
次に、色度によって輝度コントラストの影響の強さがどのように変化するかを調べた。色度が同一である刺激対ごとに、明るい色刺激に対する神経細胞集団の応答強度と暗い色刺激に対する神経細胞集団の応答強度の相関を計算した。
その結果、V4野とPITCの神経細胞集団の応答強度に対する輝度コントラストの影響は視覚刺激の色度に依存していた。V4野では青やシアンなどの色で輝度コントラストの影響が強い(図5左)のに対し、PITCでは彩度の高い色(鮮やかな色)に比べて無彩色(白/黒)や低彩度の色で輝度コントラストの強い影響がみられた(図5中央)。AITCでは、輝度コントラストの影響はどの色でもほとんどみられなかった(図5右)。
図5
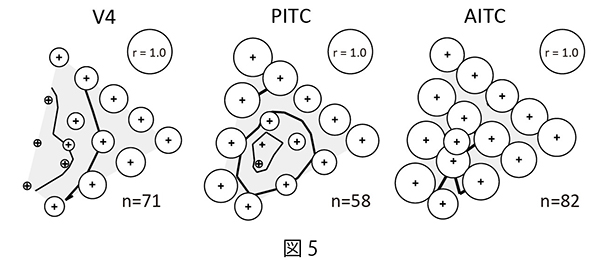
| 図5の説明 色度ごとに明るい色刺激に対する細胞集団の応答強度と暗い色刺激に対する細胞集団の応答強度の相関(r)を円のサイズで表し、色度座標の位置にプロットしている。低い相関係数(r)は、細胞集団の応答強度が明るい色刺激と暗い色刺激で異なっていたことを意味している。 |
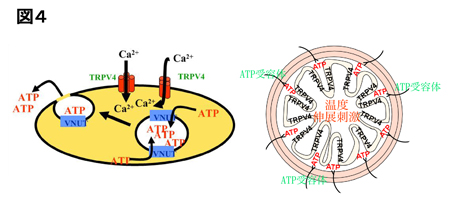
これら三領野のそれぞれの細胞集団が、背景より明るい色と暗い色の情報をどのように表現しているのかを調べた。そのために、30個の色刺激(明るい色刺激15個+暗い色刺激15個)の間で神経細胞集団の応答強度の類似関係を多次元尺度構成法(MDS)により計算し、二次元平面上に可視化した。その結果、いずれの領野においても神経細胞集団の活動は色度の情報を規則的に表現していた(図6)。さらに、V4とPITCの細胞集団の応答強度は明るい刺激セットと暗い刺激セットをはっきり区別していることが分かった(図6左、中央)。特にPITCでは、低彩度(色刺激#11)や無彩色(色刺激#16)の刺激で明るい色と暗い色の区別が顕著だった(図6中央)。一方AITCの細胞集団の活動は、輝度コントラストが異なる色刺激を区別せず、色相の順に色を規則的に表現していることが分かった(図6右)。
図6
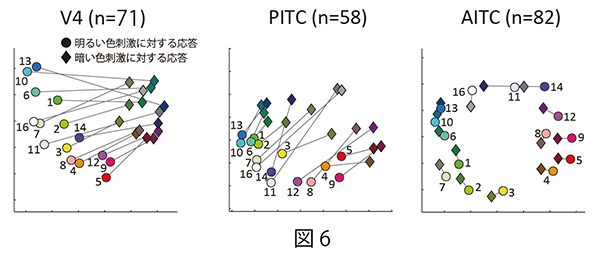
| 図6の説明 30個の色刺激に対する神経集団の応答の類似度を、相関係数rを用いて計算し、応答の非類似度(1-r)を二次元平面に投影した図。丸いシンボルが明るい色に対する神経集団応答、四角いシンボルが暗い色に対する神経集団応答を示す。シンボル間の距離が長いほど、色刺激によって引き起こされる神経集団の応答の類似度が低いことを表している。どの領野も近い色相の色が近くにプロットされており、色度の情報が規則的に表現されているが、V4 とPITCでは明るい色と暗い色が明瞭に離れてかたまっていることが分かる。 |
Namima T, Yasuda M, Banno T, Okazawa G, Komatsu H
Effects of luminance contrast on the color selectivity of neurons in the macaque area
V4 and inferior temporal cortex
The Journal of Neuroscience, November 5, 2014 • 34(45):14934–14947
]]>
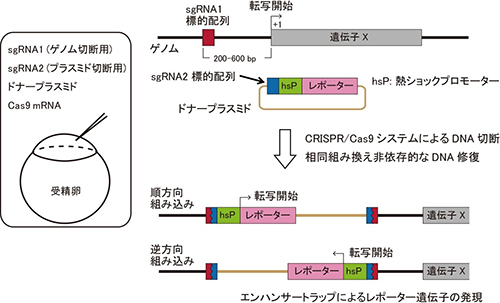 CRISPR/Cas9システムによる標的DNA配列の切断にはヌクレアーゼであるCas9と、標的配列に相同な配列を持ち、Cas9を標的配列に誘導するガイダンス分子となる短鎖RNA(sgRNA)が必要である。ノックインフィッシュ作製の際には、ゲノムを標的とするsgRNA1と、ドナープラスミドを標的とするsgRNA2、sgRNA2の標的配列を持つドナープラスミド、Cas9 mRNAを受精卵に注入した。CRISPR/Cas9システムによってドナープラスミドとゲノムが細胞内で同時に切断されると、直鎖化されたドナープラスミドがゲノム切断部位に、相同組み換え非依存的なDNA修復を介して高効率に組込まれる。本研究では、ドナープラスミドに、熱ショックプロモーターとレポーター遺伝子配列も含ませ、ゲノムのsgRNA標的配列は、標的遺伝子の上流に設計した。このドナープラスミドが標的遺伝子上流のゲノム切断部位に組込まれると、熱ショックプロモーターに、標的遺伝子の制御領域が働きかけ、標的遺伝子発現細胞におけるレポーター遺伝子の発現を促す。
CRISPR/Cas9システムによる標的DNA配列の切断にはヌクレアーゼであるCas9と、標的配列に相同な配列を持ち、Cas9を標的配列に誘導するガイダンス分子となる短鎖RNA(sgRNA)が必要である。ノックインフィッシュ作製の際には、ゲノムを標的とするsgRNA1と、ドナープラスミドを標的とするsgRNA2、sgRNA2の標的配列を持つドナープラスミド、Cas9 mRNAを受精卵に注入した。CRISPR/Cas9システムによってドナープラスミドとゲノムが細胞内で同時に切断されると、直鎖化されたドナープラスミドがゲノム切断部位に、相同組み換え非依存的なDNA修復を介して高効率に組込まれる。本研究では、ドナープラスミドに、熱ショックプロモーターとレポーター遺伝子配列も含ませ、ゲノムのsgRNA標的配列は、標的遺伝子の上流に設計した。このドナープラスミドが標的遺伝子上流のゲノム切断部位に組込まれると、熱ショックプロモーターに、標的遺伝子の制御領域が働きかけ、標的遺伝子発現細胞におけるレポーター遺伝子の発現を促す。
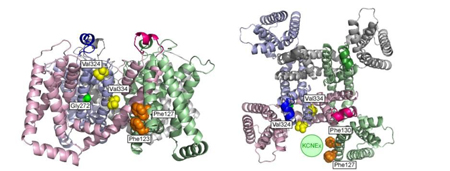
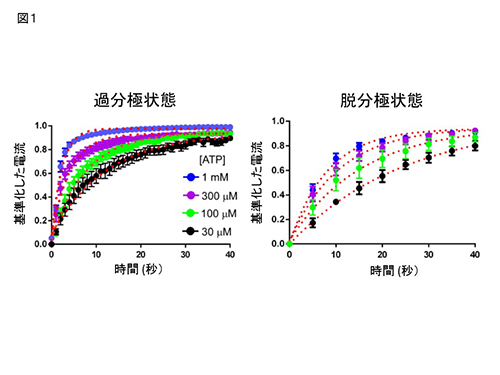 Cys変異体 (D315C & I67C) の、Cd2+ 投与による電流修飾速度は、膜電位およびATP濃度に依存して変化する。
Cys変異体 (D315C & I67C) の、Cd2+ 投与による電流修飾速度は、膜電位およびATP濃度に依存して変化する。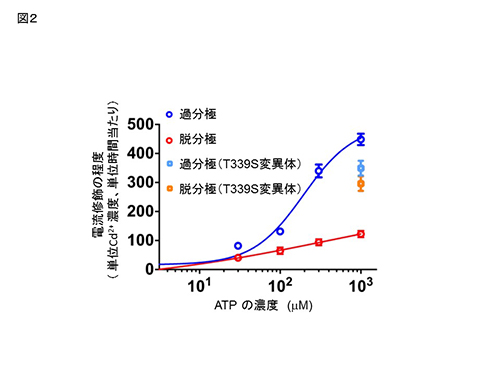 膜電位依存的ゲーティングを失ったT339S変異体 (D315C&I67C&T339S) の、Cd2+ による修飾の速度は、膜電位に依存して変化しない。青、赤は、図1で示したCys変異体 (D315C & I67C)のデータ。
膜電位依存的ゲーティングを失ったT339S変異体 (D315C&I67C&T339S) の、Cd2+ による修飾の速度は、膜電位に依存して変化しない。青、赤は、図1で示したCys変異体 (D315C & I67C)のデータ。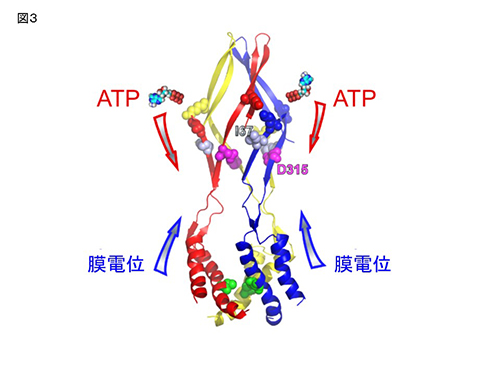 P2X2のチャネルポアとATP結合部位をつなぐリンカー領域において、ATP結合と膜電位の両方に依存して構造変化が起こる。
P2X2のチャネルポアとATP結合部位をつなぐリンカー領域において、ATP結合と膜電位の両方に依存して構造変化が起こる。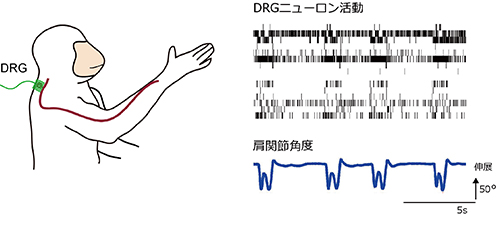 サルが到達把持運動をしているときに、16個の末梢神経が関節角度変化に対応して、それぞれの発火パターンで発火しています。
サルが到達把持運動をしているときに、16個の末梢神経が関節角度変化に対応して、それぞれの発火パターンで発火しています。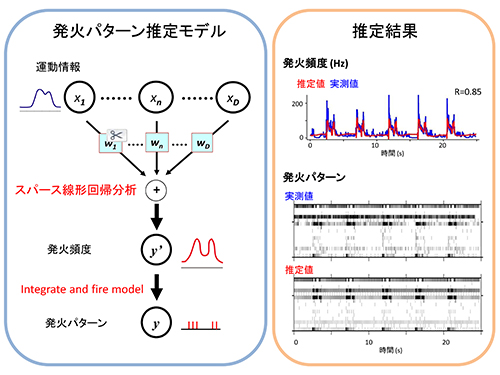 左:まず、スパース線形解析を用いて肩・肘・手首関節角度情報から末梢神経の発火頻度を計算、次に、神経発火モデル(integrate and fire model)を用いて発火頻度から発火パターンを計算しました。右上:一つの末梢神経の発火頻度の推定例。実測した発火頻度(青)と運動情報から推定した発火頻度(赤)間の相関係数(R)は0.85。右下:16個の末梢神経の実測した発火パターンと推定した発火パターン。
左:まず、スパース線形解析を用いて肩・肘・手首関節角度情報から末梢神経の発火頻度を計算、次に、神経発火モデル(integrate and fire model)を用いて発火頻度から発火パターンを計算しました。右上:一つの末梢神経の発火頻度の推定例。実測した発火頻度(青)と運動情報から推定した発火頻度(赤)間の相関係数(R)は0.85。右下:16個の末梢神経の実測した発火パターンと推定した発火パターン。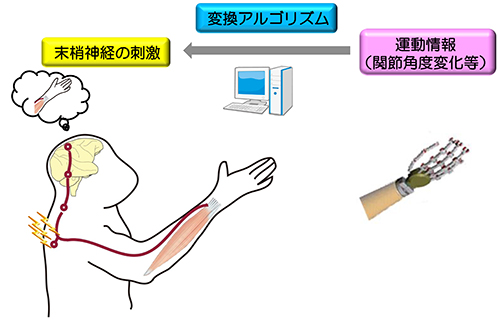 体性感覚機能を脳にフィードバックする一つの方法として、外部機器の運動情報から導き出した発火パターンで末梢神経を活動してあげることで、人工的な運動感覚を誘発することを想定しています。
体性感覚機能を脳にフィードバックする一つの方法として、外部機器の運動情報から導き出した発火パターンで末梢神経を活動してあげることで、人工的な運動感覚を誘発することを想定しています。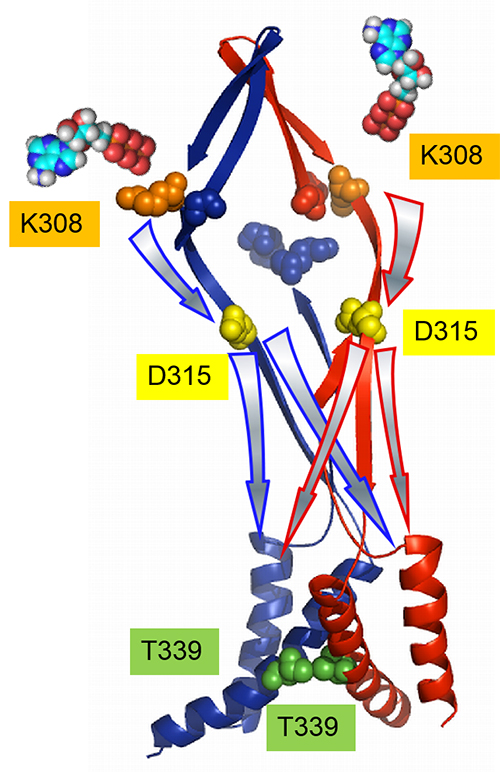 3つのサブユニットを連結したコンカテマーに、変異の位置と数をコントロールして導入することにより、2分子のATPの結合による活性化シグナルの流れを解析した。 ATPの結合のシグナル(K308) は、結合した2つのサブユニット上を、結合部位とチャネルポアをつなぐリンカー部位 (D315) まで流れる。その後、拡散し、チャネルポア部位 (T339) では、3つのサブユニットに均等に伝達される。
3つのサブユニットを連結したコンカテマーに、変異の位置と数をコントロールして導入することにより、2分子のATPの結合による活性化シグナルの流れを解析した。 ATPの結合のシグナル(K308) は、結合した2つのサブユニット上を、結合部位とチャネルポアをつなぐリンカー部位 (D315) まで流れる。その後、拡散し、チャネルポア部位 (T339) では、3つのサブユニットに均等に伝達される。
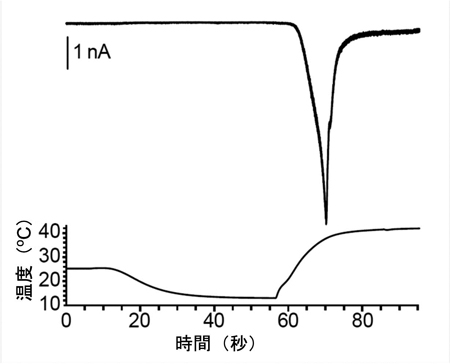
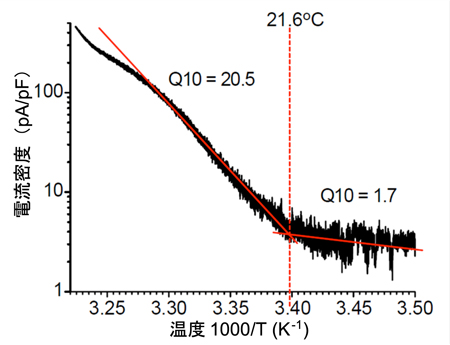 B. 熱応答電流のアレニウスプロット
B. 熱応答電流のアレニウスプロット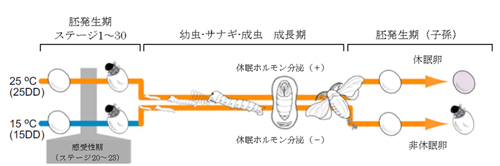 C. 胚発生期温度と休眠卵・非休眠卵表現型決定機構
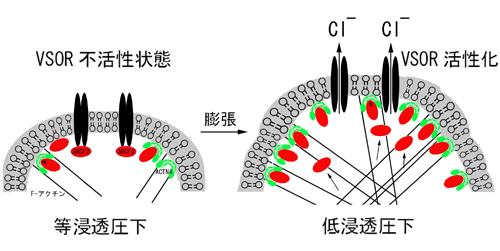
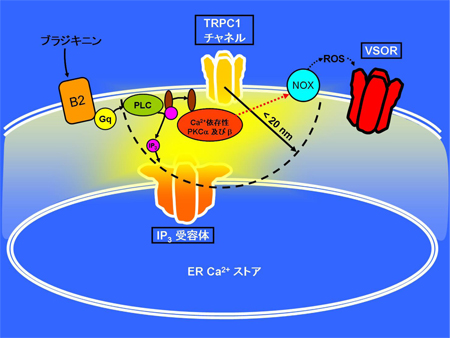
C. 胚発生期温度と休眠卵・非休眠卵表現型決定機構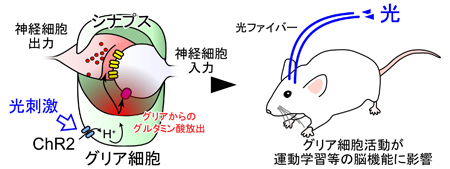 チャネルロドプシン2(ChR2)をグリア細胞に発現させ、これを光で刺激すると、グリア細胞内が酸性化することが明らかになった。この酸性化が引き金となって、細胞質内の陰イオンであるグルタミン酸が、細胞を囲む膜に存在する陰イオンチャネルを介して細胞外へと放出されることが分かった。このようにして、グリア細胞の活動は神経細胞に伝わり、最終的には学習等の脳機能に影響することが示唆された(本研究およびSasaki,…,Matsui*, PNAS, 2012 参照)。
チャネルロドプシン2(ChR2)をグリア細胞に発現させ、これを光で刺激すると、グリア細胞内が酸性化することが明らかになった。この酸性化が引き金となって、細胞質内の陰イオンであるグルタミン酸が、細胞を囲む膜に存在する陰イオンチャネルを介して細胞外へと放出されることが分かった。このようにして、グリア細胞の活動は神経細胞に伝わり、最終的には学習等の脳機能に影響することが示唆された(本研究およびSasaki,…,Matsui*, PNAS, 2012 参照)。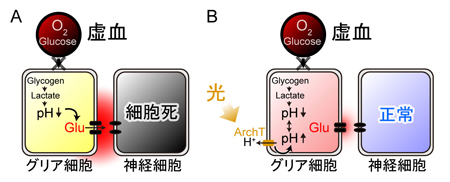 A, 脳虚血時には、血管(上丸)からの酸素とグルコースの供給が止まり、グリア細胞のグリコーゲンが分解され、乳酸が蓄積することで細胞内が酸性化する。この酸性化がグリア細胞からの過剰なグルタミン酸放出につながると考えられる。B, 脳虚血によって生じるグリア細胞内の酸性化は、グリア細胞に発現させたアーキオロドプシン(ArchT)の光刺激によって、拮抗させることができる。このとき、グリア細胞からのグルタミン酸放出が抑制され、過興奮による神経細胞死を防ぐことができた。
A, 脳虚血時には、血管(上丸)からの酸素とグルコースの供給が止まり、グリア細胞のグリコーゲンが分解され、乳酸が蓄積することで細胞内が酸性化する。この酸性化がグリア細胞からの過剰なグルタミン酸放出につながると考えられる。B, 脳虚血によって生じるグリア細胞内の酸性化は、グリア細胞に発現させたアーキオロドプシン(ArchT)の光刺激によって、拮抗させることができる。このとき、グリア細胞からのグルタミン酸放出が抑制され、過興奮による神経細胞死を防ぐことができた。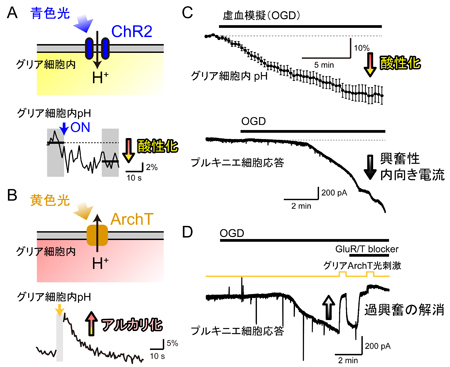 A, チャネルロドプシン2(ChR2)は、主に水素イオンを通すので、細胞内を酸性化する働きがある。細胞内pHイメージングによって酸性化を確認した。B, アーキオロドプシン(ArchT)は、逆に、水素イオンを細胞外に汲み出す光作動性のポンプである。細胞内pHはアルカリ化することが確認された。C, 小脳スライス標本を浸す灌流液から酸素とグルコースを抜く(OGD)と、グリア細胞内が急速に酸性化する。神経細胞(プルキニエ細胞)から電気記録を取ると、OGDとともに興奮性内向き電流が流れる。D, グリア細胞に発現させたArchTを光刺激してグリア細胞内の酸性化を拮抗させると、興奮性内向き電流が大幅に抑制された。グルタミン酸受容体阻害剤による効果とほぼ同じであることから、ArchT光刺激によってグリア細胞からのグルタミン酸放出が止まったと考えられる。
A, チャネルロドプシン2(ChR2)は、主に水素イオンを通すので、細胞内を酸性化する働きがある。細胞内pHイメージングによって酸性化を確認した。B, アーキオロドプシン(ArchT)は、逆に、水素イオンを細胞外に汲み出す光作動性のポンプである。細胞内pHはアルカリ化することが確認された。C, 小脳スライス標本を浸す灌流液から酸素とグルコースを抜く(OGD)と、グリア細胞内が急速に酸性化する。神経細胞(プルキニエ細胞)から電気記録を取ると、OGDとともに興奮性内向き電流が流れる。D, グリア細胞に発現させたArchTを光刺激してグリア細胞内の酸性化を拮抗させると、興奮性内向き電流が大幅に抑制された。グルタミン酸受容体阻害剤による効果とほぼ同じであることから、ArchT光刺激によってグリア細胞からのグルタミン酸放出が止まったと考えられる。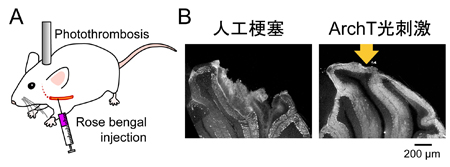 A, Rose bengal法により人工的に脳梗塞を作った。B, 3時間の局所的脳梗塞で大規模な構造的ダメージが観察されたが、その期間中、グリア細胞に発現させたArchTを間欠的に光刺激し続けた群においては、障害を抑制することができた。
A, Rose bengal法により人工的に脳梗塞を作った。B, 3時間の局所的脳梗塞で大規模な構造的ダメージが観察されたが、その期間中、グリア細胞に発現させたArchTを間欠的に光刺激し続けた群においては、障害を抑制することができた。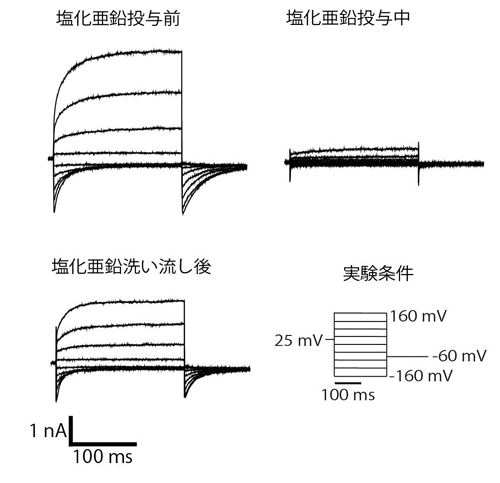 A. TRPM5チャネルを発現させたHEK293細胞にホールセルパッチクランプ法に観察されるTRPM5チャネル活性化電流
A. TRPM5チャネルを発現させたHEK293細胞にホールセルパッチクランプ法に観察されるTRPM5チャネル活性化電流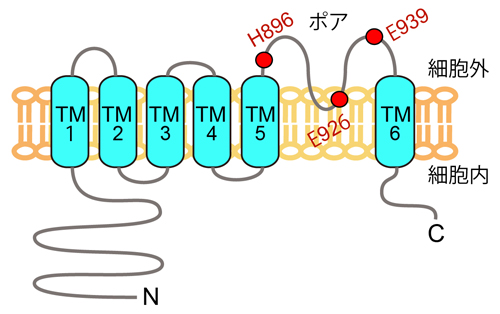 B. TRPM5チャネルの構造と亜鉛イオンの作用部位
B. TRPM5チャネルの構造と亜鉛イオンの作用部位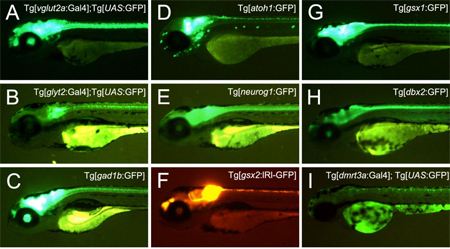
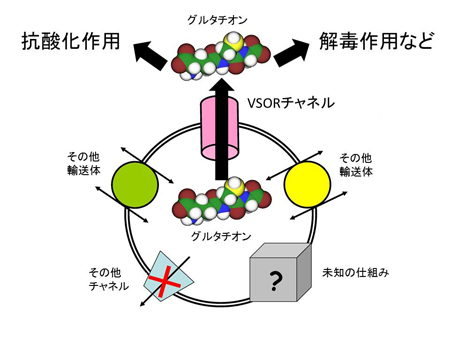
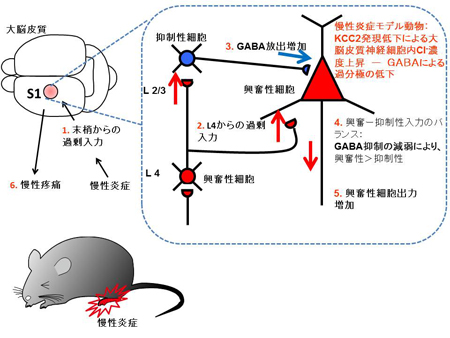
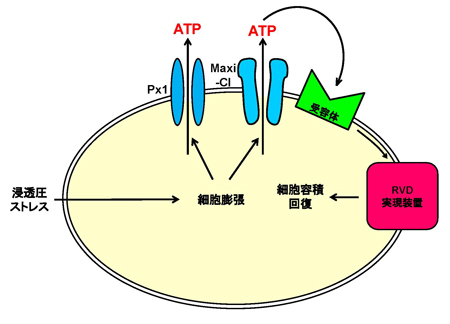
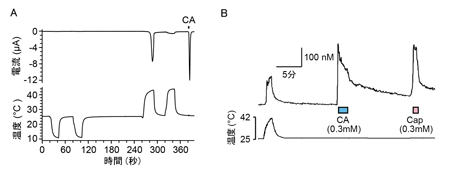
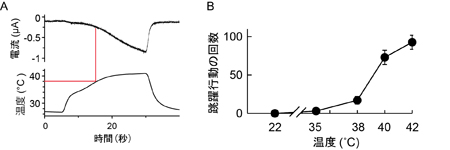
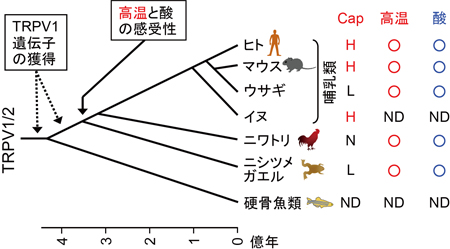
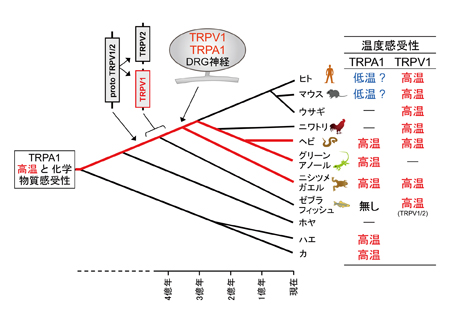 これまで解析されたTRPA1とTRPV1チャネルの特性(右表)に基づいて機能の進化過程を推定しました。TRPA1チャネルは動物の初期の進化過程で既に高温と刺激性の化学物質感受性を獲得し、その特性は脊椎動物の祖先種でも維持されていました。しかし、脊椎動物の進化過程においてTRPA1チャネルの温度に関わる機能は種間で変化しました。新たな高温センサーとしてTRPV1チャネルが脊椎動物の祖先種で獲得されたことが影響したためと考えられます。
これまで解析されたTRPA1とTRPV1チャネルの特性(右表)に基づいて機能の進化過程を推定しました。TRPA1チャネルは動物の初期の進化過程で既に高温と刺激性の化学物質感受性を獲得し、その特性は脊椎動物の祖先種でも維持されていました。しかし、脊椎動物の進化過程においてTRPA1チャネルの温度に関わる機能は種間で変化しました。新たな高温センサーとしてTRPV1チャネルが脊椎動物の祖先種で獲得されたことが影響したためと考えられます。