視覚障害者がどのように点字を「見て」いるのか―fMRI研究から判ること
抄録
視覚障害者が点字を触読するときに、視覚皮質はどのような働きを担うのだろうか。点字の触読には、単純な触知情報を語い的・意味的特性を有する言語パターンへと変換する過程が含まれる。晴眼者において、視覚による文字の同定は視覚系によって行われるが、視覚障害者による点字の知覚的処理には、体性感覚系が関与していると考えられる。近年、機能的磁気共鳴画像等の機能的神経画像法が進歩したことから、点字触読に関する神経基盤の研究が可能となった。早期視覚障害者の一次視覚皮質が点字触読に機能的に関与していることから、脳は可塑性が高く、触覚情報を視覚皮質野においても処理している可能性が示されている。
独立した感覚系
視覚および触覚情報は、目または皮膚から脳へ、物理的に独立した経路により伝達される。大脳皮質レベルにおいては、視覚と触覚とで、それぞれに情報を独自に処理する一次感覚野があり、一次視覚皮質(V1)および一次体性感覚皮質と呼ばれている。更に進んだ情報処理は、視覚情報と体性感覚情報との統合も含めて高次皮質野で起こるが、このような情報処理は、一次皮質野とは独立して行われる。V1野が損なわれることにより視覚障害が発生するが、触覚感覚の障害には至らない。同様に、一次体性感覚皮質が損なわれることによって、触覚弁別能が障害を受けるものの、視覚は影響を受けない。以上のことから、この両経路は、皮質における情報処理の最も初期段階において独立して機能していると考えられている(Pons 1996)。
点字とは何か
点字とは、よく知られているように、視覚文字を視覚障害者が触覚で知覚できるように置き換えたものであり、指でなぞって読めるように凸点による突起の組み合わせにより構成されている。点字は、点字セル(マス)と呼ばれる単位からなっており、一つの点字セルは、縦3点、横2列の6個の凸点から構成されている(図1)。6点のうちから1点以上を用いることにより、63通りの組み合わせが可能となる。一個のセルにより、アルファベット、数字、句読符や単語すら表記することができる。点字は、一つの言語であると言うよりは、英語や日本語などの言語を表記し判読するための記号と言った方がより正確であろう。点字は、ルイ・ブライユの功績によるものとされているが、この表記体系の歴史は、戦野に端を発している。19世紀初頭に、フランス軍兵士シャルル・バルビエ・ド・ラ・セールは、明かりがない戦場でも軍からの指令を読めるように、指の触覚だけで判読できる「夜間書記法」を発明した。このシステムは、12点で構成された表音記号であったが、複雑すぎたために実用化には至らなかった。1821年になって、シャルル・バルビエは、ルイ・ブライユと出会うことになるが、ブライユは4歳の頃から視力を失っていた。ルイ・ブライユは、この凸点によるシステムが優れていることに気づき、12点を6点にまで単純化した。このブライユによる点字が、世界標準となったのである。この方法は、効果的な意思疎通の方法であるばかりか、視覚障害者が文字を読めるための方法として実証されており、点字は、視覚障害者が読み書きを行ううえで、不可欠な技術となっている。
点字蝕読における行動特性
点字の触読は、複雑な認知過程であり、その中には、指の動きの制御、凸点の知覚、パターン認識および語いや意味を理解するプロセスが含まれる。
知覚
点字セルは、サイズが比較的小さいため(図1)、点字が判読できるためには、最低限の神経生理学的触覚基盤(substratum)が必要とされる。すなわち、日本語点字を触読するためには、動的二点識別閾が4mm未満であり、静的二点識別閾の場合には5mm未満でなければならない(Nakada and Dellon 1989)。
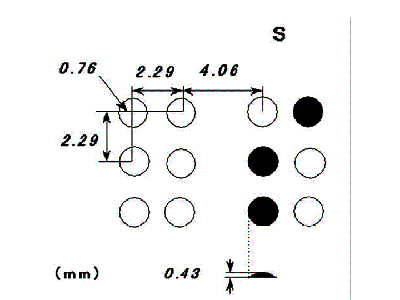
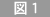 点字の構造
点字の構造
点字は、縦3点×横2列の等間隔に配置された6点(点字セルまたはマスと呼ばれる。)からなり、その組み合わせにより63通りの字形がある。各字形は、文字、数字、句読符、楽譜などを表す。
利き手
利き手は、課題の内容、経験の差、点字材料に対する慣れ度合い、片側優位性における個人差、触読習慣、文化的価値基準や触読方略に対する好み等多くの交絡因子により影響を受けると考えられる。既報文献では、点字触読の多面的な特性が示されている。すなわち、左手の方が触読能が高いとするもの(HermelinとO’Connor 1971; Rudelら1974)、右手が優れているとするもの(Fertsch 1947)、両方に差がないとするもの(Millar 1984)などさまざまである。Millar (1984) は、左利きの優位性が文字識別(それゆえ触覚情報処理過程)に関して認められ、右利きの優位性は文字特定(それゆえ語い判読過程)に認められることから、 利き手の優位性が課題内容と触読方略に関する個人的傾向の組み合わせにより決まる面があることを示唆している。この知見からは、言語処理過程と利き手には関係性のあることが示されるが、その詳細については現在まだ解明されていない。
視覚障害による影響
視覚障害により、視覚以外の感覚は、鋭敏化するのであろうか。視覚障害者において指先の触覚が優れているかという点については、以前から見解の相違がある(Axelrod 1959; Warren 1978; Hollins 1989; Pascual-LeoneとTorres 1993; Stevensら1996; Grantら2000; Van Bovenら2000)。GoldreichとKanics (2003)は、grating orientation課題(JohnsonとPhillips 1981)を完全自動化した試験系(触覚刺激の違いを検出する装置)を用いて、触覚の受動的鋭敏性を視覚障害者と晴眼者とで比較したところ、視覚障害者では触覚鋭敏性が優れていることを示す結果が得られたが、この結果は、児童期での視力、光の知覚レベルや点字判読能とは無関係であった。それゆえ、視覚障害者において触覚が鋭敏化していることに関しては、異種感覚間の可塑的変化(cross-modal plasticity)が関与している可能性が示唆される。
神経基盤との関連性
点字触読における行動特性は、神経基盤との関連性を理解することにより更によく理解することができる。ポジトロン断層画像撮影法(PET)や機能的磁気共鳴画像法(fMRI)などの脳機能画像法を用いることにより、これらの過程の神経基盤について、異種感覚間の可塑的変化と併せて研究が行われてきた(囲み記事1)。
晴眼者における触覚形状弁特性の神経基盤
触覚形状分別の神経基盤は、主として体性感覚皮質と頭頂皮質内の頭頂間溝領域前部にある(Rolandら1998; Haradaら2004)。Haradaら (2004)は、 触覚分別課題遂行時には、右背外側前頭前野、後部頭頂皮質、前補足運動野(pre-SMA)および背側運動前野(PMdr)の賦活化が非対称性に起こっており、この結果は被験者の手の左右とは無関係であったことをfMRIにより発見した。この結果は、視覚障害者による点字触読に利き手があること(Millar 1984)を部分的に説明しうると考えられる。晴眼者による触覚分別では、視覚野の関与は認められなかった(Sadatoら2002; Haradaら2004)。
視覚障害者の点字蝕読に関与した神経基盤
触覚知覚
点字触読には、単純な触知情報を語い的・意味的特性を有する言語パターンへと変換する過程が含まれる(Sadatoら1998)。視覚による文字の同定は、通常の場合視覚系が担当しているが、点字の知覚的処理には、体性感覚系が関与していると考えられる。最近の電気生理学的調査や神経画像研究からは、視覚障害者の視覚系がその本来の目的(すなわち視覚)ではなく、触知情報の処理に関与している可能性が示唆されている。[18F]フルオロ-2-デオキシグルコース(FDG)PET法を用いた研究により、Wanet-Defalqueら(1988)は、有線野および前有線野における局所脳ブドウ糖代謝率(rCMRGlc)が安静時において、触覚刺激または聴覚刺激を受けたときと同程度に上昇していることを示した。この試験においては、神経活動が課題遂行により亢進していることは示されなかったが、末梢神経系の障害による早期視覚障害者においてMRI画像が正常でありブドウ糖利用率が高いことにより、視覚障害者において、視覚野が本来の機能の代わりに視覚以外の情報処理に関与していることが示される。視覚障害者の脳波計(EEG)後頭部電極では、触覚イメージや点字触読の課題遂行に伴う賦活化が認められることから(Uhlら1991)、体性感覚入力が後頭野に送られることが考えられる。EEGよりも空間分解能が優れたO-15水PET法により、先天的視覚障害者および早期視覚障害者が点字判読その他の触覚弁別課題を遂行する際に、V1野の賦活化が認められている(Sadatoら1996; 1998)。この結果は、fMRIでも確認された(SadatoとHallett 1999; Sadatoら2002, 図2)。視覚障害者と晴眼者とでは、触覚弁別課題を実施する際には、異なる感覚器に関与した異なる神経回路が賦活化することが示されている。すなわち、二次体性感覚野(SII)において関与が認められる触覚情報処理経路は、晴眼者と比較して視覚障害者の方が賦活度が低く、その反対に、視覚形状弁別で使用される腹側後頭部領域は、視覚障害の方が賦活度が高かった(Sadatoら1998)。以上の知見からは、脳が可塑性に優れていることが示され、その可塑性により触覚情報を視覚皮質において処理することが可能となっている。
このような可塑的変化は、長期の学習による効果以上に視覚脱失に関連すると考えられが、それは、途中視覚障害者で点字が読めない被験者においても視覚連合野の賦活化が認められるからである(Sadatoら2004)。以上のことから、触知感覚の向上に関わる神経基盤は、視覚脱失による可能性が考えられる(GoldreichとKanics 2003)。
このような可塑的変化に年齢依存性のあることが、Sadatoら(2002)による最近のfMRI研究により明らかにされている。この研究では、早期視覚障害者(16歳未満で失明)および途中視覚障害者(16歳以上で失明)ともに視覚連合野が触覚弁別課題により賦活化したが、V1野の賦活化は前者の群のみで観察された。
3歳で失明した視覚障害者(左)および晴眼者(右)による点字触弁別課題遂行時の脳賦活領域。
賦活領域は、背面から撮影した高分解能MRI画像に重ね合わせた。視覚障害者では、V1を含む 後頭葉に賦活化が認められる(下左)。晴眼者では、これと対照的に、後頭葉の賦活化が見られない。
経頭蓋磁気刺激法によるV1野の機能的意義に関する同定
PETおよびfMRIにより、視覚障害者による点字触読に関与する脳神経回路が明らかにされた。しかし、脳の一領域が特定の機能に対して本質的な役割を果たしていることを証明するためには、これらの手法だけでは、不十分である。Cohenら(1997)は、 視覚障害者および晴眼者の各群に対して反復性経頭蓋磁気刺激法(rTMS)を実施した(囲み記事2)。その結果、視覚障害者の場合には、中後頭の刺激により触覚知覚に干渉が発生したが、晴眼者の場合には干渉が生じなかった。Cohenら(1997)は、更に詳細な研究を行い、この干渉が視力喪失年齢と関連性があることを示した。 すなわち、先天的視覚障害者および早期視覚障害者の中後頭にTMS刺激を加えた場合には誤答率が上昇したが、途中視覚障害者ではそのような影響が生じなかったのである。神経画像法を用いたこれまでの研究からは、早期視覚障害者のV1野は触覚弁別課題により賦活化するが、視覚連合野に関してはこのような失明年齢による影響が見られていない(Sadatoら2002)。以上のことから、早期視覚障害者のV1野は触覚弁別過程を補填するように機能的な方法により改変を起こしていることが考えられる。
関与が与えられる経路
触覚によるV1野の賦活化が想定される機序としては、視覚情報と触覚情報の入力が起こる視床が最初のレベルの処理系として考えられる。この段階までは、視覚入力と触覚入力とは完全に独立している(例えば、網膜に投影された信号は、脊髄を通って伝達する触覚情報と物理的に分断されている)。また、視床においても、視覚入力と触覚情報とは、分断された状態のままである。それゆえに、触覚刺激によるV1野の賦活化は、皮質の働きにより調節されていると思われる。視覚障害者の右手に対して触覚刺激を与えると、V1野の両側性賦活化が生じる(Pons 1996; Sadatoら1996; 1998; 2002)。この現象は、視床レベルにおける異種感覚の再構成によっては説明できない。それは、視床には対側性があること、そして、外側膝状体は同半球の皮質にしか入力信号を伝達しないからである。更に、触覚によるV1野の賦活化は、視力喪失年齢と関連性があるが、視覚連合野の賦活化には、そのような関連性が見られない。このようなことから、失明により視覚野に転送される触覚信号に関して、V1野が皮質の「エントリー・ノード」であるとは考えにくい。局所解剖学的に見て、V1野は皮質の低次のレベルにあると考えられ(すなわち、視覚情報処理の早期段階に関与するという意味において)、ボトムアップ処理において高解像度情報を受け取り、エッジ検出や局所構成過程を担っている(FellemanとVan Essen 1991)。視覚イメージ課題では、長さ、幅、向きや線の間にあるスペースの大きさ等の情報を提示するパターンの視覚化が要求されるが、この視覚イメージ課題によりV1野が機能的な方法で賦活化を受けることが示されている(Kosslynら1999)。この知見からは、イメージ処理を行う中で、脳に蓄えられた情報により、視覚野の比較的低次のレベルにおいて視覚パターンが惹起されて、形状処理が促進されると考えられる。カニクイザルの視覚野は、大部分がその他の視覚野と相互結合していて、情報を伝達した先の領域から情報を受信するようになっている(Felleman and Van Essen 1991)。そのため、視覚イメージ処理において、トップダウン信号処理が視覚連合野により調節されている可能性がある。
視覚障害者におけるトップダウン信号処理の正確な経路は不明であるものの、この活動は、多感覚領域により調節を受けていると考えられる。晴眼者においては、体性感覚入力および視覚入力は、頭頂間溝後部領域に集まるが、この領域は多感覚領域として知られている(Saitoら2003)。視覚障害者においては、視覚のボトムアップ信号処理が影響を受けているが、多感覚領域によって、触覚形状分別処理系が視覚連合野にまで拡大していることが考えられる。
視覚障害者では、このように視覚連合野の援用が起こっているために、触覚が鋭敏になっている可能性がある(GoldreichとKanics 2003)。早期視覚障害者においては、視覚イメージのトップダウン処理などにおいて、V1野も機能的な方法により援用が行われているが、途中視覚障害者ではそうではない。その結果、早期視覚障害者では形状弁別課題の成績が高くなっている(Sadatoら2002)。以上から、視覚障害者のV1野は、皮質の高次レベル、すなわち触覚刺激処理の後期に関与している皮質領域に位置すると考えられる。
意味の処理
点字触読における意味の処理に関連して、Buchelら(1998)は、先天的視覚障害者、途中視覚障害者および晴眼者の各群に対し、単語処理により生じる皮質の賦活化について、機能的神経画像法を用いて研究した。 異なる感覚(視覚、触覚)が用いられているものの、単語を提示した場合に、単語ではない文字列の場合と比較して、ブロードマン(BA)37野の賦活化が各群に共通して認められたことから、BA37野が多領域から流入する入力信号を統合する連合野であると結論付けられた。聴覚提示による動詞生成課題(Burtonら2002)および発話理解課題(Roderら2002)を用いた研究において、後頭部の賦活化が観察されており、脱失した視覚皮質が言語処理過程に関与している可能性が示されている。Amediら(2003)は、先天的視覚障害者が点字触読課題、聴覚提示による動詞生成課題および言語記憶課題を遂行する際の皮質の賦活化をfMRIにより調べたところ、視覚障害者が言語記憶課題および動詞生成課題を行う場合に、後頭部の広範囲な(V1野を含む)賦活化が認められたが、このような賦活化は、晴眼者においては見られなかった。この結果は、視覚障害者群において言語記憶能力が高かったことと関連しており、また、視覚障害者の言語記憶能力と言語記憶試験遂行時におけるV1野の賦活度との間には強い正の相関が見られたことと関連している。更に、この研究において、視覚障害者の後頭皮質には、領域特異性が認められた。すなわち、点字触読では前方領域が強く賦活化を受けるが、言語記憶および動詞生成課題では後方領域が強い賦活化を示した。rTMSを用いた試験によりAmediら(2004)は、視覚野が高次の言語処理に機能的に寄与していることを確認した。すなわち、後頭極にrTMSを印加すると、視覚障害者群では動詞生成課題の成績が低下したが、晴眼者対照群ではそのような変化は起こらなかった。また、rTMSにより言葉の意味の取り違えが広く生じたことから、脱失した視覚皮質が意味の処理過程に関与していると考えられる。
結論
視覚障害者がどのように点字を「見て」いるのか―機能的神経画像法研究から判ること
この論文は、視覚障害者による点字触読に関する一連の研究課題のうち一部を報告するものであるが、ここでは以下のような点が示されている。すなわち、技術的な面からは、ヒトの脳皮質可塑性を研究する上で、電気生理学的画像法と血行動態解析画像法とを相補的に組み合わせる手法が不可欠であることが示されており、生物学的な面では、感覚脱失により脳の可塑的再構築が起こっていることが示されたことである。V1野が視覚知覚から解放されると、触覚弁別など他の種類の感覚処理あるいは言語処理のような非感覚的情報処理に関与するように改変を受ける。このような再構築は、触覚と視覚の異種感覚統合に関わる神経回路によって調節されていると考えられることから、このような知見は、多感覚統合の理解に対しても光を投じている。
参考文献
- Amedi A, Raz N, Pianka P, Malach R, Zohary E. 2003. Early ‘visual’ cortex activation correlates with superior verbal memory performance in the blind. Nat Neurosci 6:758-66.
- Amedi A, Floel A, Knecht S, Zohary E, Cohen LG. 2004. Transcranial magnetic stimulation of the occipital pole interferes with verbal processing in blind subjects. Nat Neurosci 7:1266-70.
- Axelrod S. 1959. Effects of early blindness: performance of blind and sighted children on tactile and auditory tasks. New York: American Foundation for the Blind.
- Buchel C, Price C, Friston K. 1998. A multimodal language region in the ventral visual pathway. Nature 394:274-7.
- Burton H, Snyder AZ, Diamond JB, Raichle ME. 2002. Adaptive changes in early and late blind: a FMRI study of verb generation to heard nouns. J Neurophysiol 88:3359-71.
- Cohen LG, Weeks RA, Sadato N, Celnik P, Ishii K, Hallett M. 1999. Period of susceptibility for cross-modal plasticity in the blind. Ann Neurol 45:451-60.
- Cohen LG, Celnik P, Pascual-Leone A, Corwell B, Faiz L, Dambrosia J, Honda M, Sadato N, Gerloff C, Catala MD, Hallett M. 1997. Functional relevance of cross-modal plasticity in blind humans. Nature 389:180-3.
- Felleman DJ, Van Essen ED. 1991. Distributed hierarchical processing in the primate cerebral cortex. Cereb Cortex 1:1-47.
- Fertsch P. 1947. Hand dominance in reading Braille. Am J Psychol 60:335-49.
- Goldreich D, Kanics IM. 2003. Tactile acuity is enhanced in blindness. J Neurosci 23:3439-45.
- Grant AC, Thiagarajah MC, Sathian K. 2000. Tactile perception in blind Braille readers: a psychophysical study of acuity and hyperacuity using gratings and dot patterns. Percept Psychophys 62:301-12.
- Hallett M. 2000. Transcranial magnetic stimulation and the human brain. Nature 406:147-50.
- Harada T, Saito DN, Kashikura K, Sato T, Yonekura Y, Honda M, Sadato N. 2004. Asymmetrical neural substrates of tactile discrimination in humans: a functional magnetic resonance imaging study. J Neurosci 24:7524-30.
- Hermelin B, O’Connor N. 1971. Right and left handed reading of Braille. Nature 231:470.
- Hollins M. 1989. Understanding Blindness. Hillsdale: Erlbaum.
- Johnson KO, Phillips JR. 1981. Tactile spatial resolution. I. Two-point discrimination, gap detection, grating resolution, and letter recognition. J Neurophysiol 46:1177-92.
- Kosslyn SM, Pascual-Leone A, Felician O, Camposano S, Keenan JP, Thompson WL, Ganis G, Sukel KE, Alpert NM. 1999. The role of area 17 in visual imagery: convergent evidence from PET and rTMS. Science 284:167-70.
- Millar S. 1984. Is there a “best hand” for Braille? Cortex 20:75-87.
- Nakada M, Dellon AL. 1989. Relationship between sensibility and ability to read Braille in diabetics. Microsurgery 10:138-41.
- Ogawa S, Lee T. 1990. Magnetic resonance imaging of blood vessels at high fields: in vivo and in vitro measurements and image simulation. Magn Reson Med 16:9-18.
- Pascual-Leone A, Torres F. 1993. Plasticity of the sensorimotor cortex representation of the reading finger in Braille readers. Brain 116:39-52.
- Pons TP. 1996. Novel sensations in the congenitally blind. Nature 380:479-80.
- Raichle ME. 1987. Circulatory and metabolic correlates of brain function in normal humans. In: Handbook of Physiology, pp 643-74. Bethesda: American Physiology Society.
- Roder B, Stock O, Bien S, Neville H, Rosler F. 2002. Speech processing activates visual cortex in congenitally blind humans. Eur J Neurosci 16:930-6.
- Roland PE, O’Sullivan B, Kawashima R. 1998. Shape and roughness activate different somatosensory areas in the human brain. Proc Natl Acad Sci USA 95:3295-300.
- Rudel R, Denckla M, Spalten E. 1974. The functional asymmetry of Braille letter learning in normal sighted children. Neuology 24:733-8.
- Sadato N, Hallett M. 1999. fMRI occipital activation by tactile stimulation in a blind man. Neurology 52:423.
- Sadato N, Okada T, Honda M, Yonekura Y. 2002. Critical period for cross-modal plasticity in blind humans: a functional MRI study. Neuroimage 16:389-400.
- Sadato N, Okada T, Kubota K, Yonekura Y. 2004. Tactile discrimination activates the visual cortex of the recently blind naive to Braille: a functional magnetic resonance imaging study in humans. Neurosci Lett 359:49-52.
- Sadato N, Pascual-Leone A, Grafman J, Deiber MP, Ibanez V, Hallett M. 1998. Neural networks for Braille reading by the blind. Brain 121:1213-29.
- Sadato N, Pascual-Leone A, Grafman J, Ibanez V, Deiber M-P, Dold G, Hallett M. 1996. Activation of the primary visual cortex by Braille reading in blind subjects. Nature 380:526-8.
- Saito DN, Okada T, Morita Y, Yonekura Y, Sadato N. 2003. Tactile-visual cross-modal shape matching: a functional MRI study. Cog Brain Res 17:14-25.
- Stevens JC, Foulke E, Patterson MQ. 1996. Tactile acuity, aging, and Braille reading in long-term blindness. J Exp Psychol Hum Percept Perform Appl 2:91-106.
- Uhl F, Franzen P, Lindinger G, Lang W, Deecke L. 1991. On the functionality of the visually deprived occipital cortex in early blind person. Neurosci Lett 124:256-9.
- Van Boven RW, Hamilton RH, Kauffman T, Keenan JP, Pascual-Leone A. 2000. Tactile spatial resolution in blind Braille readers. Neurology 54:2230-6.
- Wanet-Defalque M-C, Veraart C, Volder AD, Metz R, Michel C, Dooms G, Goffinet A. 1988. High metabolic activity in the visual cortex of early blind human subjects. Brain Res 446:369-73.
- Warren DH. 1978. Perception by the blind. In: Handbook of Perception (Carterette EC, Friedman MP, eds), pp 65-90. New York: Academic Press.