4統合生理研究系
4.1感覚運動調節研究部門
高次脳機能(顔認知、音楽認知、言語認知など)に関連する脳反応、各種感覚(視覚、聴覚、体性感覚、痛覚、嗅覚)に対する脳反応、運動に関連する脳反応などを、各種ニューロイメージング手法(脳波、脳磁図、機能的MRI、近赤外線分光法、経頭蓋磁気刺激)を用いて研究している。2007年に発表した論文のうち代表的な3編を紹介する。
Ogino Y, Nemoto H, Inui K, Saito S, Kakigi R, Goto F (2007) Inner experience of pain: imagination of pain while viewing images showing painful events forms subjective pain representation in human brain. Cerebral Cortex 17:1139-1146.
以前から「心の痛み」といった表現が良く使われてきたが、その実態は不明であった。機能的MRI (fMRI)を用いて、痛みを想像したときの脳活動を計測したところ、それは本当に痛みを与えられたときとほぼ同一の場所であった(図1)。また、恐怖の画像を呈示した時には扁桃体に活動が見られ、同じような不快な画像に対しても、心が痛い時とは異なる脳活動が見られた。本研究により、確かに「心は痛む」ことを、最新の脳科学機能画像を用いて初めて科学的に証明することに成功した。
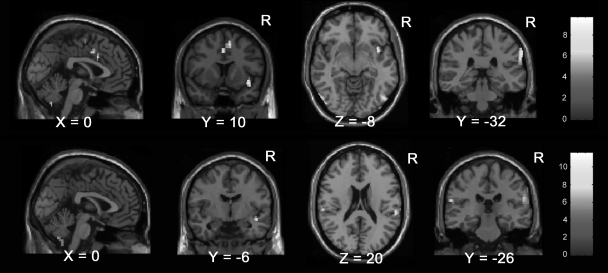
図. 心の痛みを感じたときの脳活動:帯状回前部と両側半球の島に特異的な活動が見られ、それは実際に針などで痛みを与えたときとほぼ同様の部位であった。
Miki K, Watanabe S, Honda Y, Nakamura M, Kakigi R (2007) Effects of face contour and features on early occipitotemporal activity when viewing eye movement. NeuroImage 35:1624-1635.
被験者に運動が知覚される仮現運動を利用した4条件の視覚刺激を用いて誘発脳磁場を測定した。(1) CDL: 模式的な顔の絵(輪郭、目、口)の中で目の部分が動く刺激。(2) CD: CDLより口を取り除いた刺激。(3) DL: CDLより輪郭を取り除いた刺激。(4) D: CDLより輪郭と口を除いた刺激。D条件では単なる2つの点の動きにしか見えないが、他の条件、特にCDL条件では顔に見えるため、目の動きとして認知される。脳磁場反応の活動の大きさは、CDL条件時に有意に大きかった。動きそのもの自体は条件間で違いがないにもかかわらず、条件によって活動の大きさに差がみられた。このことより、ヒトのMT/V5野では、運動そのものに対する反応に加えて、「目の動き」に対する特異的な活動が起こり、その際、顔の輪郭とパーツの情報が重要な役割を担っている可能性が示唆された。
Okamoto H, Stracke H, Ross B, Kakigi R, Pantev C (2007) Left hemispheric dominance during auditory processing in noisy environment. BMC Biol. 5, 52. (Online Journal) http://www.biomedcentral.com/content/pdf/1741-7007-5-52.pdf
日常生活において私達は、自分達にとって意味のある音信号と同時に、自分達にとって意味のない雑音に晒されている。音信号処理の過程で、これら音信号と雑音によって引き起こされた神経活動は互いに干渉しあうと考えられる。今回の実験では脳磁図を用いて、雑音と信号音によって引き起こされる神経活動を計測した。音信号と雑音の相互干渉作用により、音信号により惹起された脳神経活動の有意な減少がみられたが、左半球の神経活動の低下は右半球に比し、有意に小さかった。この結果より、騒音環境下においては、左半球の方が右半球より音信号処理に重要な働きをすると考えられる。今回私達が発見した、騒音環境下における左半球の優位性は、声のような複雑な音声刺激処理における、左半球優位性に繋がるのではないか、と考えられる。(ドイツ ミュンスター大学・カナダ トロント大学との共同研究)。
4.2生体システム研究部門
本研究部門は、「随意運動の脳内機序の解明」を主要な課題として、大脳皮質・大脳基底核・小脳・脳幹などを中心に、手指を精緻に動かすような随意性の高い運動から、歩行や咀嚼のように半ば自動化されたものまで、幅広い運動の脳内機構について、包括的に明らかにするという目的で、主に霊長類を用い神経生理学的手法、あるいは神経生理学的手法と神経解剖学的手法を組み合わせて研究を行っている。また、これらが障害を受けた際の疾患の病態、さらに治療法のメカニズムについての研究も進めている。霊長類を用いた研究に加えて、様々な疾患モデルを扱えるという利点を生かして、げっ歯類を用いた神経生理学的研究も行っている。
2007年に発表した論文を紹介する。
Kita H, Chiken S, Tachibana Y, Nambu A (2007) Serotonin modulates pallidal neuronal activity in the awake monkey. J Neurosci 27:75-83.
縫線核のセロトニン作動性ニューロンは、淡蒼球内節・外節に投射しており、また淡蒼球にはセロトニン受容体が存在する。セロトニンが大脳基底核の運動機能に関わっていると考えられているが、淡蒼球におけるセロトニンの生理学的機能については不明なことが多い。他の脳部位におけるこれまでの神経解剖学的検索や脳スライス実験によって、以下のような可能性が考えられる。
- セロトニンは、シナプス前膜にある5-HT1B受容体を介して、GABA作動性の抑制を減弱させる。
- セロトニンは、シナプス後膜にある5-HT1A受容体を介して、ニューロンの発射頻度を減少させる。
- セロトニンは、シナプス後膜にある5-HT2C、5-HT4、5-HT7受容体を介して、ニューロンを興奮させる。
上記の可能性を確かめるため、覚醒下サルを用い、局所にセロトニンの作動薬や拮抗薬を注入し、淡蒼球ニューロンの自発発射や大脳皮質運動野を電気刺激した際の反応に与える効果について調べた。このようなin vivo実験では、受容体のタイプや作動部位の決定までは難しいが、以下のような可能性が示唆された。
- セロトニンは、5-HT1B受容体を介して、GABA作動性の抑制を強く減弱した。
- この減弱は、淡蒼球外節では、それ以外の受容体や機序も関与している可能性がある。
- セロトニンは、5-HT1A受容体を介して、グルタミン酸作動性の興奮作用を減弱させた。
- 本実験においては、セロトニンのニューロンに対する強い直接作用は観察できなかった。
これらの実験結果は、セロトニンが淡蒼球内節・外節へのシナプス伝達を変化させることにより、運動制御にも関わっていることを示唆するものである。
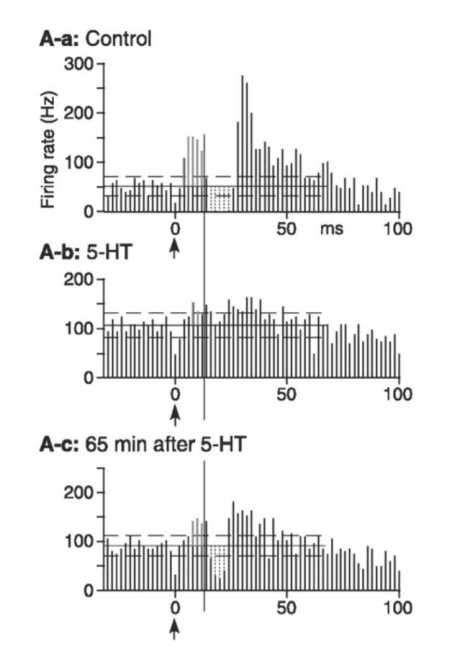
図の説明
大脳皮質運動野刺激(矢印)に対する淡蒼球外節ニューロンの反応を、刺激後時間ヒストグラムで示す。セロトニン(5-HT)の局所投与により、皮質由来の興奮と抑制が減弱していることがわかる。