5大脳皮質機能研究系
5.1脳形態解析研究部門
脳形態解析部門では、脳の神経細胞膜上の受容体やイオンチャネルなどの機能分子の機能と局在や動態を電気生理学的方法と免疫電子顕微鏡法などによって定量的に明らかにし、神経伝達調節メカニズムを研究してきた。具体的な研究内容をいくつか列挙する。
1)シナプス内グルタミン酸受容体局在とシナプス応答との関係: これまでのAMPA受容体局在解析から、AMPA受容体のシナプス内局在にはほぼ均一にシナプス全体に分布するパターンとモザイク状に分布するパターンの2つがあり、このパターンが入力線維の種類に依存して形成されることがわかって来た。例えば外側膝状体の中継細胞に形成される2種のシナプス(視神経-中継細胞シナプスと大脳皮質-中継細胞シナプス)のうち前者は全体分布型、後者はモザイク型であることが分かった。そこで観察されたシナプス内で起こるグルタミン酸素量放出とAMPA受容体応答のシミュレーションを行うことでシナプス応答を推定したところ、どちらのシナプスでも受容体数の増加に比例した安定したシナプス応答が得られることが予想された。この結果は個々のシナプスは、受容体配置にかかわらず受容体数の変化が忠実に応答強度に反映されるように形成されていることを示唆している。
2)海馬における長期増強現象とグルタミン酸受容体の密度変化: シナプス伝達の長期的な機能変化は学習・記憶の細胞レベルの素過程と考えられており、その分子メカニズムについて内外で精力的に研究されているが、個々のシナプス膜上の機能分子の局在や量がどの様に変化するのかは、技術的制約により殆ど明らかになっていない。そこで定量的な細胞膜上分子の局在解析が可能なSDS凍結割断レプリカ標識法を導入し、神経伝達物質受容体の局在を個々のシナプスレベルで解析できるように改良した。この方法を用いて長期増強現象を誘導したところ、シナプス内AMPA受容体密度が増加し、NMDA受容体密度が減少する事が明らかとなり、また、シナプス外細胞膜上に存在するAMPA受容体がシナプスへ組み込まれるAMPA受容体の供給元になっていることが示唆された。また、シナプスが形成される樹状突起スパインとシナプスのサイズ及び受容体局在との関係を解析し、シナプス機能の増強に伴いスパインの形態変化と受容体増加及びシナプス面積増加が短時間で起き、受容体増加にはGluR1サブユニットを含む受容体が関与することを示す結果を得た。一方、動物が探索行動を行う事により上記LTP実験で得られた受容体レベルの変化が起きることも確認した。
3)シナプス-グリア複合環境の動的変化による情報伝達制御: シナプス前終末部から放出された伝達物質は細胞外空間を拡散し、その広がり方に従って、神経細胞間の情報伝達の特性は決定される(Matsui & von Gersdorff, 2006)。 伝達物質の拡散を制御し、学習や記憶に重要とされるシナプス辺縁の受容体の活性化を制御できる格好の位置に、グリア細胞が存在する。我々は、シナプス-グリア複合環境の動的変化が、伝達物質濃度の時空間特性にどう影響するのか調べている。これまで、シナプス前細胞からグリア細胞のほうに向けて異所性のシナプス小胞放出があり、これがニューロン-グリア間の素早い情報伝達を担っていることを示してきた(Matsui& Jahr, 2006)。この情報伝達によってグリア細胞の形態や機能が制御されている可能性を、二光子励起イメージングによって解析している。グリア細胞によるシナプスの包囲率の相違が、シナプス伝達にどんな影響を与えるのかを、電気生理学・電子顕微鏡法も組み合わせて解明する。
4)前脳基底核のシナプス伝達機構と大脳基底核シナプス再生機構: 前脳基底核は中枢アセチルコリン性ニューロンの起始核であり、記憶、学習、注意等の生理的機能と密接に関係するとともに、その病的状態としてアルツハイマー病との関連が示唆されている。アセチルコリン性ニューロンへの興奮性および抑制性シナプス伝達機構および修飾機構の生後発達変化につき、ニューロン同定の新たな手法を導入しつつ、電気生理学的解析および形態学的解析を行ない、アセチルコリン性ニューロンへのグルタミン酸遊離制御におけるドーパミンD1型受容体とP/Q型カルシウムチャネルとの選択的共役を明らかにした。現在その生後発達変化について検討を加えている。
大脳基底核シナプスおよび神経回路の再生機構の詳細は不明である。現在、実験的に脳虚血状態を起こしたラットおよびパーキンソン病モデルラットを用いて、傷害された線条体神経細胞、シナプス再生経過および再生機構を明らかにする目的で、形態学的および電気生理学的解析を行なっている。本プロジェクトによる基礎的データが、脳梗塞等の疾患に対する新たな治療法開発につながることを期待したい。
5.2大脳神経回路論研究部門
大脳機能を支える局所神経回路の構成を調べることを目標にし、これまでに大脳皮質のニューロンタイプを、軸索投射・発火・物質発現のパターンから同定してきた。現在は、同定してきた構成要素(投射・介在ニューロンサブタイプ)から皮質回路がどのような原則で組み上げられているかを明らかにすることを目指している。今年度は、(1)錐体細胞スパインへの興奮・抑制入力の関係、(2)アセチルコリンの新皮質及び海馬錐体細胞への作用を解析した。
アセチルコリンの錐体細胞への作用を、新皮質の層ごとに、海馬内の領域ごとに比較した。新皮質では領野によらず、一過性のアセチルコリン投与に対して5層のものは著明に過分極するのに対して、2・3層のものでは電位変化はみられなかった
(図1)。 アセチルコリンを海馬錐体細胞の細胞体付近に一過性に与えたところ、CA3領域では過分極が見られなかったが、CA1領域では、M1型ムスカリン受容体、細胞内カルシウム上昇を介したSKチャネルによる過分極が起きた
(図2)。新皮質では2・3層から5層錐体細胞への結合が主要な興奮性経路の一つなのに対して、海馬ではCA3からCA1錐体細胞へのシナプスが主な興奮経路である。皮質外への投射については、それぞれ5層とCA1錐体細胞が担っている。従って、アセチルコリンは、層構造下位の興奮性細胞の発火を一過性に抑制し、皮質外への出力を遮断する可能性がある。アセチルコリン作用・皮質内経路・皮質外投射様式を合わせて考えると、新皮質2・3層錐体細胞はCA3錐体細胞に、5層のはCA1のものに対応すると考えられた。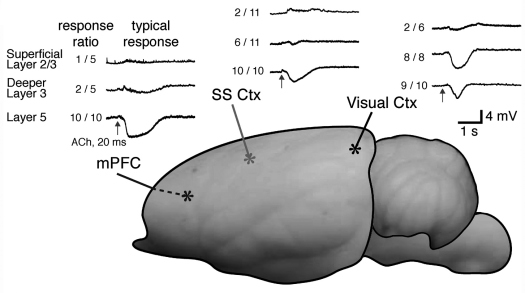
図1. 複数の皮質領域での一過性アセチルコリン応答の比較。どの領野でも深層の錐体細胞だけにSKチャネルによる過分極応答が見られた。
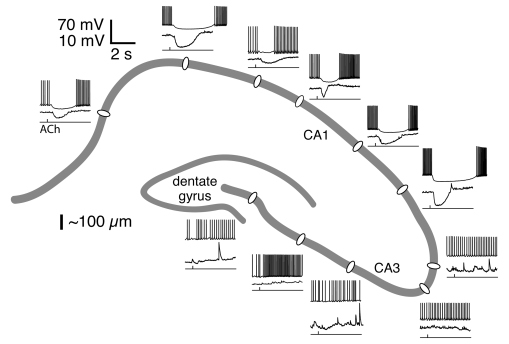
図2.アセチルコリンは海馬CA1の錐体細胞を一過性に過分極させ、スパイク発火時にはそれを抑制した。この過分極・発火抑制はCA3錐体細胞にはみられなかった。この反応性は、CA1やCA3領域内での細胞体位置には依存していなかった。
錐体細胞への興奮性入力の多くは樹状突起のスパインにシナプスを作る。スパインへの興奮・抑制性入力パターンを電顕で解析した。生理的・化学的・形態的に同定した非錐体細胞軸索のシナプス標的構造を超薄切片像から3次元的に再構築して調べたところ、複数のGABA細胞サブタイプの軸索がスパインにシナプスを作り、その同じスパインに興奮性終末がみられた。視床と皮質からの終末をそれぞれの特異的な抗体で同定し、そのシナプス後構造を超薄切片像から再構築すると、視床入力を受けるスパインへの一部に同時にGABA作働性入力があることがわかった。非錐体細胞の複数のサブタイプが、視床からのスパインへの興奮性入力の一部を選択的に抑えることを明らかにできた(図3)。
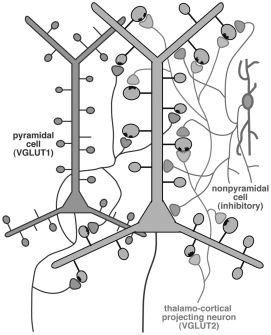
図3. 皮質錐体細胞由来の軸索終末には主にVGLUT1が、視床からのものにはVGLUT2が発現している。多様な皮質細胞のサブタイプが錐体細胞のスパインにGABA作動性終末をつくるが、同じスパインへの興奮性入力はVGLUT2陽性であった。
5.3心理生理学研究部門
認知,記憶,思考,行動,情動,社会能力などに関連する脳活動を中心に,ヒトを対象とした実験的研究を推進している。脳神経活動に伴う局所的な循環やエネルギー代謝の変化をとらえる脳機能イメーシング(機能的MRI)と,時間分解能にすぐれた電気生理学的手法を統合的にもちいることにより,高次脳機能を動的かつ大局的に理解することを目指している。機能局在と機能連関のダイナミックな変化を画像化することにより,感覚脱失に伴う神経活動の変化や発達および学習による新たな機能の獲得など,高次脳機能の可塑性のメカニズムに迫る一方、近年は特に 自己と他者との関係(社会的認知)にかかわる神経基盤に重点を置いて研究している。
2007年に発表した論文のうち代表的な1編を紹介する。
Morita T, Itakura S, Saito DN, Nakashita S, Harada T, Kochiyama T, Sadato N (in press) The Role of the Right Prefrontal Cortex in Self-Evaluation of the Face: A Functional Magnetic Resonance Imaging Study. J Cogn Neurosci.
ヒトは1歳半頃になると,鏡に映る自分の顔を見てそれが自分であることに気付く.いわゆる自己顔認知ができるようになる.それに対して,自分で自分についての評価をおこない,その評価に伴った自己意識感情(例:恥ずかしさ,罪悪感)が生起するのは2-3歳ごろといわれている.自己顔の認知プロセスには右側前頭-頭頂ネットワークが重要であることが示されてきたものの自己顔の評価に関わる脳領域については明らかにされていない.そこで,自己顔認知と自己顔評価に関わる神経基盤の違いを明らかにすることを目的として機能的MRIを19名の被験者に対して行った.ビデオ画像から切り出された不自然な顔写真を用い,自己顔および他者顔を評価しているときの脳活動をfMRIを用いて計測した.自己顔に対する写真写り評価の低下に伴い,恥ずかしさの増大が見られたのに対して,他者顔についてはそのような関係は見られなかった.他者顔を評価している時に比べて,自己顔を評価している時に,右側前頭前野,両側の島,帯状回および両側の後頭葉の活動が有意に増加していた.右側前頭前野の活動のピークは,運動前野付近および,より前方の下前頭回付近にそれぞれ見られ,これら2領域の活動は異なる様相を示していた.運動前野の活動量は,公的意識尺度の強度に伴い変化していた.この結果は右側運動前野が自己顔認知に関連していることを示唆するものである.また,この領域はミラーシステムの一部ともいわれており,他者顔の観察時に賦活することが知られており、自己顔認知とミラーシステムの密接な関連を裏付ける結果といえる.一方,右側下前頭回は,自己顔評価時のみ有意な活動が見られた.また,この領域の活動量は,自己顔に対する恥ずかしさの低下に伴い活動が増加していた.これらの結果より,右側の下前頭回は,基準となる自己像との関連性に基づく自己評価において重要な役割を果たしていると考えられる.