3 生体情報研究系
3.1 感覚認知情報研究部門
感覚認知情報部門は視知覚および視覚認知の神経機構を研究対象としている。主に無麻酔のサルの視覚野に微小電極を刺入してニューロン活動を記録し、ニューロンの刺激選択性や、異なる種類の刺激への反応の分布を調べることにより、視覚情報の脳内表現を明らかにすることを試みると共に、さまざまな行動課題時のニューロン活動を分析することにより、それらの視覚情報が知覚や行動にどのように関係しているかを調べている。これに関しては具体的な課題として(1)初期視覚野における輪郭とその折れ曲がりの表現、(2)下側頭皮質における色選択性ニューロンが色知覚や色弁別にどのように関わっているか、(3)高次視覚野における色情報処理経路の同定、(4)視覚関連領野における要素的な刺激のグルーピングのメカニズムなどに関する研究を行った。またサルでfMRIを用いた実験により、(5)色に選択的に表現する脳領域の同定を勧めている。
2008年に上記の(2)について発表した論文を紹介する。
Matsumora T, Koida K and Komatsu H Relationship between color discrimination and neural responses in the inferior temporal cortex of the monkey. J Neurophysiol (in press).
色覚の成立には大脳皮質が不可欠である。これはヒトで大脳腹側の領域が損傷された時に色の知覚が失われる症状(大脳性色盲)の存在から明らかである。それでは大脳皮質のどの部分のニューロンがどのように働いて色覚が成立するのだろうか?サルの大脳皮質では下側頭皮質の損傷により、色弁別の永続的な障害が起きることが破壊実験により示されており、下側頭皮質が色知覚に極めて重要な役割を果たすものと考えられる。下側頭皮質には色選択性を示すニューロンが存在し、それらは下側頭皮質の一部の場所に集まって存在することを我々はこれまでに示してきた。しかしそれらのニューロンが色知覚とどのように関係しているかを調べた研究はこれまでに行なわれていなかった。そこで、我々は色弁別課題を訓練したサルの下側頭皮質からニューロン活動の記録を行ない、ニューロン活動とサルの色弁別行動の関係を定量的に調べることにより、下側頭皮質ニューロンが色知覚にどのように寄与しているかを明らかにすることを試みた。
色弁別課題では色度図上で等間隔でならんだ7色(サンプル色セット、#1から#7)のうちの一つが呈示され、サルはその色がサンプル色セットの両端のどちらにより近い色であるかを答える。各サンプル色に対して#7側と答えた割合を求めて描いたグラフは心理測定関数(psychometric function)と呼ばれ、この関数から色弁別閾値を求めることができる。一方サルが色弁別課題を行っている時に記録されたニューロン活動から、サンプル色ごとに#7側を選ぶ割合をROC解析により求めて神経測定関数を作成し、これからニューロン活動にもとづく色弁別閾値を求めた。同時に記録したサルの色弁別閾値とニューロン活動から求めた色弁別閾値を比較した。6つのニューロンについて比較した結果を図に示している。ニューロンの方がサル全体より高い感度をもつもの(a, d)、同じ程度の感度のもの(b, e)、低いもの(c, f)などさまざまなものが存在する。gは神経測定関数と心理測定関数から求めた弁別閾値の関係を示すが、平均としてニューロンの弁別閾値はサルの弁別閾値より1.5倍大きく、ニューロンの方が感度がやや低かった。行動から求めた色弁別閾値とニューロン活動から求めた色弁別閾値の色度図上での変動には高い正の相関が見られ、TE野の色選択性ニューロンの活動と色知覚には強い関係があることが示された。また各試行におけるニューロン活動とその試行でサルが行った行動の間にも有意な相関が見られた。しかし試行毎の相関の強さとニューロンの感度には全く相関が見られなかった。この結果は少数の感度の高いニューロンだけではなく、多数のニューロンの弱い信号が合計されたものがサルの色知覚や色弁別行動に用いられていることを示唆している。
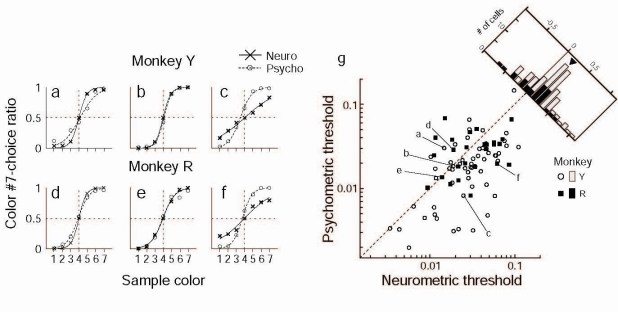
図 : aからfは6つのニューロンについて心理測定関数と神経測定関数を比較した結果を示す。ニューロンの方がサル全体より高い感度をもつもの(a,
d)、同じ程度の感度のもの(b, e)、低いもの(c, f)などさまざまである。gは神経測定関数と心理測定関数から求めた弁別閾値の関係を示すが、平均としてニューロンの弁別閾値はサルの弁別閾値より1.5倍大きかった
3.2 神経シグナル研究部門
神経シグナル部門では、神経回路機能の理解をめざしてボトムアップ的な研究を行っている。 主な研究手法は、脳スライスを用いた電気生理学であるが、遺伝子改変マウスを用いた行動解析や、生体マウス脳からのシングルユニット記録等も行なっている。具体的なターゲットは、視床、大脳皮質、小脳、海馬など幅広い領域にわたるが、基本的方針として、分子の細部に入っていくのではなく、個体との関係においてできるだけ生理学的意義のある現象を追うことにしている。
Ca2+/カルモジュリン依存性プロテインキナーゼⅡによる学習・記憶の制御
Ca2+/カルモジュリン依存性プロテインキナーゼII(CaMKII)は、学習・記憶に重要な働きをする。 その主要サブユニットCaMKIIαの不活性型ノックインマウス(K42R)を作製し、機能解析を進めている。 K42Rホモマウスでは、海馬CA1領域におけるシナプスの長期増強(LTP)が障害されているのみならず、海馬LTPと相関が深い受動的回避テストでも顕著な障害が観察された。CaMKIIは非常に量の多いタンパクであることから、構造タンパク的役割が重要であるとの説が提唱されているが、本研究より酵素活性は学習・記憶に不可欠であることが示された。現在より詳細な行動実験を行い、学習・記憶におけるCaMKIIの役割を分析している。
小脳顆粒細胞-介在ニューロン間興奮性シナプス伝達のペアパルス増強
小脳スライス‐パッチクランプ法を用いて、顆粒細胞軸索(上向性線維)の電気刺激に伴い分子層介在ニューロンから記録される興奮性シナプス後電流(EPSC)のペアパルス増強を調べた。2発目EPSCは、振幅値のみならず減衰時定数が著しく増大した。薬理学的検討ならびにキネティクス解析を行い、振幅増大は放出確率ならびに放出多重性の増強により、減衰時間増大は遅延した非同期的シナプス小胞放出の増強により生じることを示唆する結果を得た。 また放出多重性と非同期放出にはCaV2.1(P/Q型)Ca2+チャネルが、振幅増大にはCaV2.2(N型)が深く関与していることが明らかとなった。 減衰時定数の増大は、高頻度の興奮性入力の条件で脱分極を強く促進する。 混在するCa2+チャネルサブタイプの機能な違いはCalyx of Heldの巨大シナプス終末で知られているが、通常の大きさのシナプスでの実証は初めてである。
カイニン酸受容体による皮質視床シナプス伝達のシナプス前修飾
皮質視床線維(CT)は視床投射細胞(TC)のみならず視床網様体核細胞(RT)にも投射している。カイニン酸受容体が、CT-TCシナプスとCT-RTシナプスのシナプス伝達に対し、相反する作用があることを明らかにした。すなわち、カイニン酸受容体の活性化がCT-TCシナプスでは神経伝達物質の放出を抑え、CT-RTシナプスでは神経伝達物質の放出を促進することを、paired-plus ratio、coefficient of variation、MK801による放出確率への影響、素量解析などであきらかにした。さらに、カイニン酸受容体のサブユニットであるGluR5がCT-TCシナプス前性修飾に関わっていることも明らかになった。またこれらのカイニン酸受容体を介したこのような作用は、CTの高頻度刺激による内因性グルタミン酸によっても引き起こされることが明らかになった。Miyata M, Imoto K. J Physiol (in press).
神経修飾物質によるシナプス統合のスイッチング
視床から大脳皮質への信号伝達は興奮性であるが、抑制回路がその伝達を制御している。 スライスパッチクランプクランプ法とダイナミッククランプ法を組み合わせ、 視床-大脳皮質間のシナプス統合を調べた結果、2つの入力がある場合に、単一の神経回路配線がタイミングに依存する三種類のシナプス統合を呈することを示した。またシナプス統合は、異なる神経修飾物質によってスイッチされること、を明らかにした。 さらに大脳皮質抑制細胞の一種であるFast-Spiking (FS)細胞におけるEPSPの加算特性が、そのシナプス統合の成立に非常に重要であることを見出した。
3.3 神経分化研究部門
神経分化研究部門は、体制が比較的単純な脊椎動物であるゼブラフィッシュを用いて、脊髄神経回路の発生機構および回路機能の解析を行っている。胚期、幼生期初期には、ゼブラフィッシュの体はほぼ透明である。この利点を生かし、蛍光タンパク質を特定のクラスの神経細胞に発現させ、それら神経細胞を生きたまま可視化することを研究手法の中心に据えて研究を進めている。以下に2008年度に発表した論文の概要を記す。
Kimura Y, Satou C & Higashijima S (2008) V2a and V2b neurons are generated by the final divisions of pair-producing progenitors in the zebrafish spinal cord. Development 135:3001-3005.
脊髄腹側のp2前駆体細胞領域からは二種類の介在神経細胞、V2aとV2bニューロンが生じることが知られている。これまでに、V2aとV2bの分化には、分裂終了後の細胞間で起こるDelta-Notchシグナリングを介した相互作用が関与することが示されていた。しかし、V2aとV2bの細胞系譜は不明であり、上記の細胞間相互作用がどの細胞どうしで起こるかについては分かっていなかった。我々はp2前駆体細胞の最終分裂直前にGFPを発現するトランスジェニックゼブラフィッシュTg[vsx1:GFP]を作成し、この問題に取り組んだ。経時観察により、GFPでラベルされたp2前駆体細胞の大部分は、一度だけ分裂し、V2aとV2bニューロンを対で生むことが分かった。この結果は、V2aとV2bニューロンが一対のニューロンを生じる分裂、かつ非対称な分裂によって生じることを示している。さらに、Delta-Notchシグナリングの相互作用が姉妹細胞間で起こることが、この分裂が結果として非対称になるために重要な役割を果たしていることを示唆する結果も得られた。この細胞運命決定のメカニズムはショウジョウバエの神経母細胞が、非対称分裂によって二つの異なるニューロンを生じる場合と良く似ている。しかし、ショウジョウバエの場合と異なり、ゼブラフィッシュのp2前駆体細胞の分裂軸の方向は決まっていないことも分かった。これらの結果は、脊椎動物の神経発生において、一対のニューロンを生じる神経前駆体細胞の最終分裂が、分裂軸の方向性に依存しないメカニズムを介して、二つの異なるニューロンを生むことができることを示している。
本研究は、脊椎動物の中枢神経系において、1つの神経前駆体細胞から非対称分裂により2つの異なるタイプのニューロンが再現的に生じることを示す初めての例である。哺乳類脳形成においても、1つの前駆体細胞によりニューロンペアを産生する例が知られている。そこにおいても、本研究で示されたように、Delta-Notchを介した姉妹細胞同士の相互作用により2つの異なるタイプのニューロンが生じる機構が存在するかもしれない。
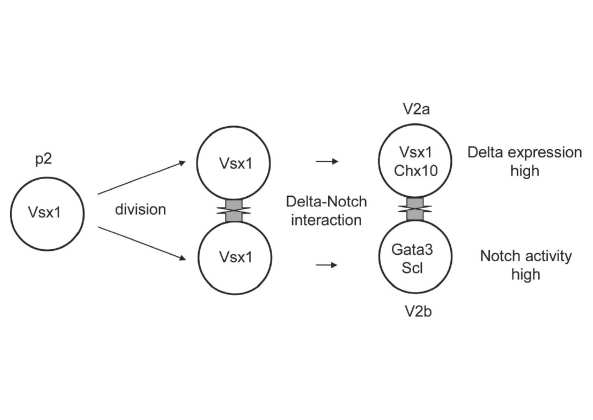
図1. V2ニューロン分化の模式図
この模式図は、ニューロンペアー産生p2前駆体細胞からの1つの細胞系譜を示す。Vsx1を発現するp2前駆体細胞は最終分裂を行う。子孫の姉妹細胞間でDelta-Notchを介した細胞間相互作用が起こり、片方の細胞で高いdeltaの発現が起こり、もう片方の細胞ではNotchの活性が強くなる。最終的に、前者はV2aニューロンへ分化し、後者はV2bニューロンへ分化する。Chx10、Gata3/SclはV2aニューロン、V2b
ニューロンのマーカー遺伝子。