3 生体情報研究系
3.1 感覚認知情報研究部門
感覚認知情報部門は視知覚および視覚認知の神経機構を研究対象としている。主に無麻酔のサルの視覚野に微小電極を刺入してニューロン活動を記録し、ニューロンの刺激選択性や、異なる種類の刺激への反応の分布を調べることにより、視覚情報の脳内表現を明らかにすることを試みると共に、さまざまな行動課題時のニューロン活動を分析することにより、それらの視覚情報が知覚や行動にどのように関係しているかを調べている。具体的な課題として(1)初期視覚野における輪郭とその折れ曲がりの表現、(2)下側頭皮質における色選択性ニューロンが色知覚や色弁別にどのように関わっているか、(3)高次視覚野における色情報処理経路の同定、(4)視覚関連領野における要素的な刺激のグルーピングのメカニズムに関する研究、(5)質感に関わる視覚情報の脳内表現の研究などを行った。またサルでfMRIを用いた実験により、(6)色に選択的に表現する脳領域の同定を進めその成果を論文として発表した。これは生理研のMRI装置を用いた初めてのサルのfMRI研究の発表である。2009年に上記の(3)について発表した論文を紹介する。
Yasuda M, Banno T, Komatsu H (2009) Color selectivity of neurons in the
posterior inferior temporal cortex of the macaque monkey. Cerebral Cortex,
doi:10.1093/cercor/bhp227.
サルの下側頭皮質は大脳皮質の腹側に存在する高次領野で、損傷されると色の識別が障害されることからヒトで色知覚に重要な役割を果たす腹側高次視覚領野と対応するものと考えられる。我々は下側頭皮質後部(PIT)でニューロンの刺激選択特性を詳細にマッピングした結果、鋭い色選択性を持つ細胞が密集して存在し、視野の場所を表現する地図を持つ領域が存在することを発見しPITC(下側頭皮質後部色領域)と名づけた。この領域は色情報処理に深く関係しているものと考えられる。
図5.3Aは今回マッピングを行ったPITの場所を示す。この場所は下側頭皮質の入口にあたる場所である。図5.3Bは脳の写真の上に我々が新しく発見したPITCの位置を示している。PITCは後中側頭溝(PMTS)をまたがって存在し、上部のニューロンは中心視野に受容野を持つが、下部に移動すると受容野は周辺視野を含むようになり、更に後部では上視野、前部では下視野に受容野を持つという、全体として大ざっぱな視野の地図を持っていた。大脳視覚野にはいくつもの視野の地図が存在するが、別々の視野地図は別々の機能に対応すると考えられている。従ってPITCも特定の機能に関係した一つの領野に対応するものと考えられる。我々はCIE-xy色度図で一定間隔に分布した同じ明るさ(輝度)の色刺激のセットを使って、ニューロンの反応を調べた。その結果PITC内から記録されたニューロンの多くは鋭い色選択性を持っていたが、PITCの外で記録されたニューロンは鋭い選択性を示さなかった。鋭い色選択性を持つニューロンが多いことから、PITCは色情報処理に深く関係した領野であると推測される。
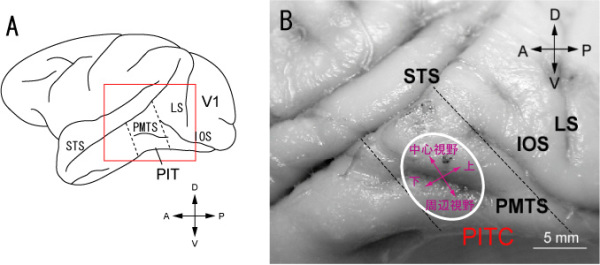
図5.3. A, サルの脳の外側面とPITの場所. B, PIT付近の脳の拡大写真とPITCの場所および視野表現.
図5.4はPITCとその周りの領域のニューロンの性質をマッピングした結果を示している。図2Aは色選択性の鋭さを示している。SとBはそれぞれ一定の定量的な基準を上回る鋭い色選択性を示した細胞(S)と示さなかった細胞(B)を表す。点線より下の領域に鋭い色選択性を持つニューロンが密集して存在していた。この部分をPITCと名づけた。図2Bは受容野が視野のどこに位置していたかを示している。PITCの上の方には中心視野に受容野を持つ細胞(F)が多いのに対し、下の方ではより周辺視野を含む受容野を持ち、更に上視野に受容野を持つ細胞(○)が後に存在し、下視野に受容野を持つ細胞(●)が前の方に存在し、全体として大ざっぱな視野地図を持っていることが分かった。この領域のニューロン活動が色知覚の成立とどのように関わっているのかを知ることが今後の課題である。
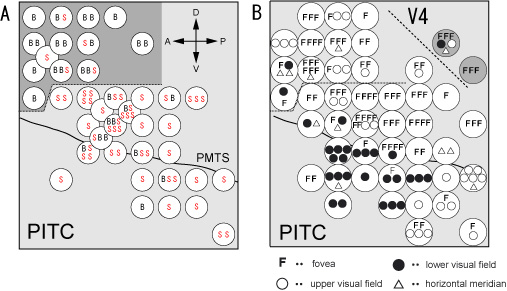
図5.4. A, PITC付近から記録したニューロンの色選択性の分布. Sは鋭い色選択性を示した細胞。Bは広い色選択性を示した細胞 (sparseness
index0.3で区分). B, PITC付近から記録したニューロンの受容野の視野位置の分布.
3.2 神経シグナル研究部門
神経シグナル部門では、神経回路機能の理解をめざしてボトムアップ的な研究を行っている。 主な研究手法は、脳スライスを用いた電気生理学であるが、遺伝子改変マウスを用いた行動解析や、生体マウス脳からのシングルユニット記録等も行なっている。今年は脊髄のin vivoパッチクランプ記録の開発者である古江秀昌准教授のグループが加わり、研究のターゲットも拡大した。
Ca2+/カルモジュリン依存性プロテインキナーゼⅡによる学習・記憶の制御
Ca2+/カルモジュリン依存性プロテインキナーゼII(CaMKII)は、学習・記憶に重要な働きをする。 その主要サブユニットCaMKIIαの不活性型ノックインマウス(K42R)を作製し、機能解析を進めている (Yamagata et al., J Neurosci 2009)。 K42Rホモマウスは、CaMKIIαノックアウトマウスよりも顕著な海馬依存性の記憶障害があるが、扁桃体等他の部位が関与すると考えられる記憶の形成・消去過程を、行動実験により検討している。
小脳顆粒細胞-介在ニューロン間興奮性シナプス伝達のペアパルス増強
小脳スライス‐パッチクランプ法を用いて、顆粒細胞軸索(上向性線維)の電気刺激に伴い分子層介在ニューロンから記録される興奮性シナプス後電流 (EPSC)のペアパルス増強を調べた。2発目EPSCは、振幅値のみならず減衰時定数が著しく増大した。この増大には、シナプス小胞の多重放出が関与しているが、それだけでは説明できないため、グリアによるグルタミン酸処理機構も含めて検討中である。
欠神発作における大脳基底核の役割
欠神発作モデルマウスであるtotteringマウスを用いて、大脳基底核がてんかん発作でどのような働きをするかを検討している。視床下核におけるシナプス伝達を遮断すると欠神発作はほとんど消失する。一方、脳スライス標本を用いたパッチクランプ記録より、視床下核神経細胞のHyperpolarization-activated (Ih) channelの活性が、低下していることが示された。薬理学的および深部脳刺激による視床下核神経細胞の活性調節が、てんかん発作にどのような影響を与えるかを検討中である。
脊髄における痛覚処理機構
これまで脊髄の痛覚伝達・処理機構に関わる神経細胞の電気生理学的測定を行ってきたが、これらの細胞群がどのようにグループ分けされ、どのような機能を果たしてるかに関しては不明な点が多く残されている。この問題点を解決するためには、電気生理学だけではなく、形態学を含めた神経回路の理解が必須である。現在、神経細胞の形態、各種抗体による染色性などの検討を行っている。
また骨のがんによる疼痛は、モルヒネ抵抗性であることなどより、他の疼痛とは異なる性質を持つことが知られている。大腿骨に骨肉腫細胞を注入して作成した骨がんモデルマウスを用いて、脊髄の痛覚伝達に関与するシナプスの特性を検討している。脊髄の多セグメントに渡ってシナプスの変化が検出されている。
さらに、脊髄の痛覚処理機構は、脳幹から下向性の抑制を受けることが知られている。この下向性抑制の一部はノルアドレナリン性であり青斑核に由来することから、in vivoで青斑核の活動を測定することを目指している。脳幹部分は拍動などのためにin vivoパッチクランプを行うのは困難なぶいであるが、いろいろな測定条件を試みて青斑核からの記録方法の開発を行っている。この実験条件では、薬剤を青斑核等の脳幹神経細胞に直接投与できるため、麻酔薬等の作用機序の解析に有用であると期待されている。
これらの研究の一部はすでに投稿準備中であり、他の研究に関しても順次論文としてまとめていく予定である。
3.3 神経分化研究部門
吉村教授を中心とする研究グループでは、大脳皮質視覚野の神経回路特性とその経験依存的発達メカニズムの解析を行っている。本年度は、脳スライス標本を用いたパッチクランプ記録装置の立ち上げを行い、視覚野2/3層の錐体細胞への特異的神経結合が、生後の視覚体験に依存して形成されるかについて検討した。生後直後からの暗室飼育により視覚体験を経ていないラットの視覚野よりスライス標本を作成し、ケージドグルタミン酸による光スキャン局所刺激法と、複数の2/3層錐体細胞からの同時ホールセル記録法を用いてその神経回路を解析した。その結果、暗室飼育したラット視覚野においては、2/3層錐体細胞が形成する興奮性結合の検出確率と強度は、正常な視覚体験を経たラットの視覚野と比較して有意に低下していた。また、正常な視覚野においては、興奮性シナプスで結合している2/3層錐体細胞ペアは、その周辺の興奮性細胞からの入力を高い頻度で共有し、非常に微細なスケールの特異的神経回路網を視覚野内に形成しているが、暗室飼育した視覚野においては、2/3層錐体細胞間の興奮性結合の有無に関わらず、どのペアにおいても周辺細胞からの共通入力は稀であった。以上の結果は、微小神経回路網の形成には、遺伝的機構のみならず生後の正常な視覚入力に依存して神経結合が精緻化される過程が必要であることを示唆する。
東島准教授を中心とするグループは、体制が比較的単純な脊椎動物であるゼブラフィッシュを用いて、脊髄神経回路の発生機構および回路機能の解析を行っている。胚期、幼生期初期には、ゼブラフィッシュの体はほぼ透明である。この利点を生かし、蛍光タンパク質を特定のクラスの神経細胞に発現させ、それら神経細胞を生きたまま可視化することを研究手法の中心に据えて研究を進めている。以下に2009年度に発表した論文(Satou, et al., 2009)の概要を記す。硬骨魚類の早い逃避行動には、後脳に存在するマウスナー細胞が重要な役割を果たしている。マウスナー細胞は脊髄内で運動ニューロンにシナプス結合しており、マウスナー細胞の活動電位は体の屈曲を引き起こす。運動ニューロンに加え、マウスナー細胞は、あるクラスの交差型抑制性介在ニューロンに電気シナプスを作っていることが知られていた。しかし、それら交差型抑制性介在ニューロンの役割は明らかではなかった。本研究では、当該交差型抑制性介在ニューロン(以下、CoLoニューロンとよぶ)が特異的にGFPでラベルされたエンハンサートラップラインを用い、CoLoニューロンを、解剖学的、電気生理学的、行動学的に詳細に解析した。その結果、CoLoニューロンは逃避行動の方向決定において重要な役割を果たしていることを明らかにした。これは、脊髄レベルでの情報処理が、行動の最終アウトプットに非常に大きな役割を果たしうることを示すものであり、中枢神経系の情報処理の機構に対して重要な視点を与えるものである。