1 分子生理研究系
1.1 神経機能素子研究部門
神経機能素子研究部門では、イオンチャネル、受容体、G 蛋白質等の構造と機能に関する研究を展開している。具体的には、 (1) ATP受容体チャネルの、膜電位とATP濃度に依存するゲーティングの分子機構、(2) Gタンパク質結合型受容体の動的構造変化とシグナリングの多様性、(3) KCNQ1-KCNE1 チャネル複合体の会合ストイキオメトリーと機能調節機構、(4) Kv1.2チャネルの細胞外K+に依存する脱活性化遅延の構造基盤と分子機構、(5) マウスとヒトの TRPA1チャネルのカフェインに対する応答の相違の一次構造の基盤の同定、(6) 小脳 lobule10のプルキンエ細胞の、GABAB受容体活性化によって開くCs+透過性を持つK+チャネルの分子同定、(7) 下垂体隆起葉に発現するOrphan代謝型受容体Prrt3の機能解析と遺伝子破壊マウスの作成、等を主たる研究目標とし学際的アプローチにより研究を進めている。2010年の発表論文のうち代表的なもの Nakajo K, Ulbrich MH, Kubo Y, Isacoff EY (2010) Stoichiometry of the KCNQ1-KCNE1 ion channel complex. Proc Natl Acad Sci USA 107: 18862-18867. の内容を以下に紹介する。
KCNQ1 は電位依存性K+チャネルのαサブユニットをコードする遺伝子であり、心臓をはじめさまざまな臓器で発現しており、不整脈の原因遺伝子としても知られている。心臓ではKCNE1 とよばれる1回膜貫通型の膜タンパク質と複合体を構成しており、IKs とよばれる非常にゆっくりと活性化する電流を担っている。KCNQ1 は他の電位依存性K+チャネルと同様4量体であることはほぼ間違いないが、4量体のKCNQ1 に対して何個のKCNE1 が結合するかについては未だ決着がついていなかった。それはひとつには、これまでの解析がマクロ電流を元になされたものであり、全体の平均像を見ていたにすぎないからだと考えられる。そこで、我々は、全反射蛍光顕微鏡を用いた一分子レベルのイメージングを用い、蛍光分子の一分子レベルの退色数を数えることにより(図)、一つのKCNQ1-KCNE1チャネル複合体に何個のKCNE1 が含まれているかを解析した。その結果、4つのKCNQ1 サブユニット(1個のイオンチャネル)に対し、1-4個のKCNE1 サブユニットが含まれていることが明らかになった。すなわち、4:1から4:4までさまざまなストイキオメトリーをもつチャネル複合体が存在することが示された。さらに、KCNQ1 とKCNE1 の相対的な発現密度を変えることで、ストイキオメトリーの分布が変化することを見出した。以上の結果により、1つのKCNQ1 チャネル複合体に含まれるKCNE1 のサブユニット数は固定されたものではなく、状況依存的に変化しうることが示唆された。
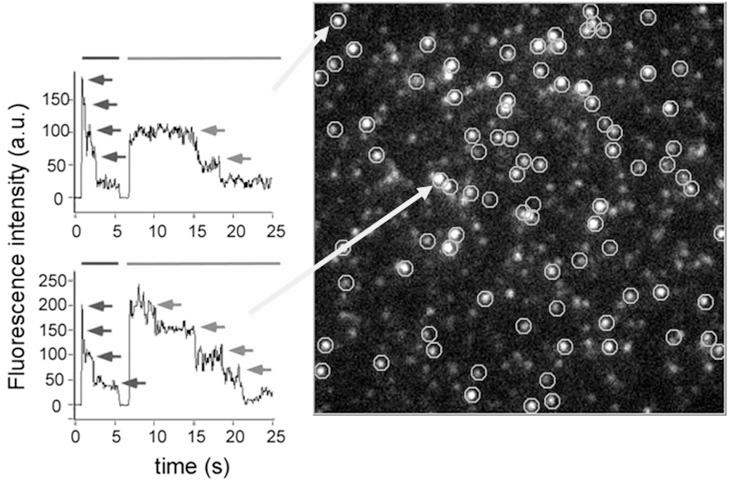
図の説明:(右)緑色蛍光タンパク質GFPを付加したKCNE1分子の一分子蛍光像。円で囲まれたスポットのひとつひとつがKCNQ1-KCNE1複合体を示す。(左)記録の後半が、矢印で示された各スポットから観察されたGFPの退色イベントを示す。上の例は2分子、下の例では4分子のGFPすなわちKCNE1分子の存在が確認できる。記録の前半は、KCNQ1につないだ赤色蛍光タンパク質の退色イベントで、4量体であることが示されている。
1.2 分子神経生理研究部門
概要
分子神経生理部門では哺乳類神経系の発生・分化、特に神経上皮細胞(神経幹細胞)からどのようにして全く機能の異なる細胞種(神経細胞、アストロサイト、オリゴデンドロサイトなど)が分化してくるのか、について研究を進めている。また、得られた新しい概念や技術は臨床研究への応用を視野に入れながら、病態の解析にも努力している。
脳神経系では他の組織とは異なり多様性が大である。そのため、他組織の分化研究とは異なり、細胞株や脳細胞の分散培養系を用いた研究ではその本質に迫るには限界がある。われわれはin vitro で得られた結果を絶えずin vivo に戻して解析するだけでなく、神経系の細胞系譜の解析や移動様式の解析をも精力的に行っている。
近年、成人脳内にも神経幹細胞が存在し、神経細胞を再生する能力を有することが明らかとなった。この成人における神経幹細胞数の維持機構についても研究している。
糖蛋白質糖鎖解析法を開発し極めて微量な試料からの構造解析が可能となった。脳内において、新しい糖鎖構造を発見し、その生理学的意義について検討している。
1)グリア細胞の発生・分化:
中枢神経系において、神経細胞の多様性は発生期に形成されることが知られている。その一方でグリア細胞の発生メカニズム、発生起源、及び発生期に規定付けられる機能的多様性については未だ不明な点が多い。本研究室ではグリア細胞の発生を規定する分子としてOlig2転写因子に着目し、グリア細胞の発生・分化機構の解明を試みている。また、前脳オリゴデンドロサイトの背側と腹側の境界領域に位置する発生起源についても解析を進めるとともに、小脳オリゴデンドロサイトの発生起源に関しても解析を行っている。更に、アストロサイトにも発生する場所に応じてサブタイプが存在する可能性を想定し、脊髄をモデルとした解析系の確立をニワトリ胚を用いて行っている。
2)神経幹細胞の生成と維持:
神経幹細胞は全ての神経細胞・グリア細胞の供給源であり、脳の構築に非常に重要であるにもかかわらず、その生成の分子機構は不明な点が多い。本研究室は早期胚のepiblastにおいて神経幹細胞の前駆細胞である未分化神経幹細胞の培養に成功し、神経幹細胞の誘導にNotchシグナルの活性化が必須であることを解明した。さらに、Notchシグナルの活性化の初期段階をglial cells missing 1/2遺伝子が担っていることを明らかにし、詳細な分子機構の解明を進めている。そこで得られた知見をES細胞に適応し、試験管内でのES細胞から神経幹細胞の誘導を試みる。一方、神経幹細胞は成体脳においても一部の領域(海馬や嗅球など)に新生神経細胞を供給し、脳機能維持に必要であることが示唆された。特に、海馬における神経新生は、記憶や学習といった脳の高次機能と関係する可能性が指摘されている。本研究室では、躁うつ病の治療に用いられる気分安定薬が成体脳における神経幹細胞の自己複製能を高めること、それがNotchシグナルの活性化によることを明らかにした。今後は、気分安定薬のNotchシグナルにおける分子標的の同定や、神経幹細胞の増加が気分を安定させるそのメカニズムの解明に取組む。
3)神経細胞の移動・軸索ガイダンス:
脊髄の組織構築形成をモデルとして、回路網形成と細胞移動の制御機構を解析するために、脊髄の回路網形成および視床網様核の細胞の移動について研究を行った。今までに、一次求心性線維の脊髄内における回路網形成においては、脊髄背外側部で一過性に発現するnetrin 1が抑制的に作用してwaiting periodを形成することを明らかにした。また、感覚神経の軸索ガイダンスについては、Olig2ノックアウトマウスで感覚ニューロンの軸索ガイダンスの異常がみられ、それが運動ニューロンの欠損によると推測され、詳しい解析を進めている。腹側視床の形成については、発生中の視床網様核において、Zona limitans intrathamica周囲のOlig2陽性細胞から分化したニューロンの局在が、発生が進むにつれて脳室側から外側方向(軟膜側)へと変化していた。このことは、発生中の腹側視床ではニューロンが脳室側から軟膜側へ移動して視床網様核を形成している可能性が考えられ、経時観察による解析を検討している。
4)グリア細胞の機能と病態:
グリア細胞の重要な機能の一つにシナプス伝達の調節がある。近年、グルタミン酸とATPがアストロサイトから放出され、シナプス伝達を調節することが提唱されているが、その放出部位、放出頻度など不明な点が多い。名古屋大学(現 東京大学)廣瀬教授の開発したプローブを用いて培養アストロサイトから放出されるグルタミン酸を可視化することに成功した。また名古屋大学 曽我部教授のグループと共同で、ルシフェリン発光を利用し、培養アストロサイトから放出されるATPの可視化にも成功した。これらのイメージングの解析から、ATP刺激によるグルタミン酸放出、グルタミン酸刺激によるATP放出が確認されたが、これらの放出は培養アストロサイトの1〜10%の細胞だけで観察されること、その放出時間が数十秒あることなどが分かった。今後は、放出に関わる分子の同定、放出の観察されるアストロサイトの特徴抽出、ATPとグルタミン酸との同時測定が課題となった。
またアストロサイトからのATP放出異常マウスを見出し解析したところ、抑制系の選択的な抑制によりLTPが誘導され易くなっていることが分かった。慢性脱髄巣におけるオリゴデンドロサイト成熟阻害因子を探索したところ、シスタチンFが見つかった。シスタチンFはミクログリアが髄鞘膜を貪食すると発現誘導され、髄鞘再生が抑制されると発現が消失する興味深い因子である。今後シスタチンFの遺伝子発現を制御できるマウスを作製する。
5)N-結合型糖鎖の構造決定と機能解析:
糖鎖を有する分子は細胞表面や細胞外に存在し、細胞間相互作用やシグナル伝達に深く関わっている。これまでに我々は (1)脳内糖鎖発現パターンが発生時期に劇的に変化すること、(2)いくつかの糖鎖の発現量が顕著に変化すること、(3)シアル酸付加糖鎖の構造解析から、大脳皮質の発達過程において劇的に変化する新規シアル酸糖鎖が存在することを明らかにした。本年度はHPLCを使ったN-結合型糖鎖の微量解析法を開発し、この解析法を用いて生検資料や精製した中枢神経や末梢神経由来の髄鞘の糖鎖構造を決定した。また、ゲルから単一糖蛋白質を切り出し、高回収率で糖鎖を解析する技術を開発した。
また本方法を利用して、血清中の肝癌マーカーを見出した。我々はN-結合型糖鎖解析過程でLewis X糖鎖構造の合成に関わる新規フコース転移酵素FUT10を見出した。そして、糖蛋白質糖鎖にフコースを転移することや神経細胞移動に関与することを明らかにした。
6)多機能遺伝子改変システム: FAST systemの構築と応用:
ある特定の遺伝子の機能を知る、特定の遺伝子異常が原因となる遺伝性疾患のモデル動物を作成するといった目的には、遺伝子過剰発現、ノックアウトが最低必要になる。場合によっては時期特異的、細胞種特異的な操作も要求される。これらの要求に応えるためには、それぞれ別個にマウスを作成する必要があった。今回新たに構築した多機能遺伝子改変システム: FAST systemは、1回のジーンターゲッティングで、5種類の異なる遺伝子操作、単純ノックアウト、レスキュー、異所性発現、過剰発現、可逆的ノックアウトが可能になる。このシステムを用いて、アストロサイト特異的遺伝性疾患であるMegalencephalic leukoencephalopathy with subcortical cystsのモデルマウスを樹立した。更に、このシステムを応用して光感受性蛋白、チャネルロドプシンを細胞種特異的に発現させることに成功し、主に線条体投射神経とグリア細胞の光操作を開始した。
1.3 ナノ形態生理研究部門
1. 位相差電子顕微鏡の開発と医学生物学への応用(永山G)
国際拠点化研究、総研大学融合研究、民間共同研究、一般共同研究および計画研究等々において位相差電子顕微鏡の開発と生物学・医学応用を行った。手法開発としては、位相板無帯電化研究、位相差トモグラフィー解析法高度化、位相差電顕単粒子解析法高度化を行った。生物学・医学応用としては、複合たんぱく質、ウィルス、核酸脂質複合系、膜たんぱく質リポーム複合系、シアノバクテリア、血小板、培養細胞などを用いた研究を行った。
国際拠点化研究では膜電位の位相差電顕によるイメージング法を開発するためYale大学のとの国際共同研究を始めた。総研大学融合研究では糖尿病治療のための膵島移植におけるインシュリン細胞死の問題を統合イメージング手法で研究開始した。特に蛍光顕微鏡と電顕との相関観察に力を入れた。民間共同研究では、位相差法の高度化の最大の難関、位相板帯電問題の解決のための研究開発を行い、上質の位相差像を可能とする無帯電化位相板が実現できるようになった。各種共同研究では、i) モーター蛋白質の単粒子解析(J Str Biol)、ⅱ) ウィルスの単粒子解析(Structure, J Virology)、ⅲ)核酸脂質複合体の立体構造解析などを行った。iv) 位相差法における画像歪み(位相板孔由来のfringing問題)の理論解析、v)電顕・光顕ハイブリッド顕微鏡の生物応用として、蛍光たんぱく質融合アクチン発現細胞を用いた性能検証、vi) トモグラフィーにおける情報欠損問題の解決法研究などを行った。
2. 外部分泌腺の形態生理学(村上G)
i) 傍細胞輸送の形態学的生理学的基盤:2009年につづき、傍細胞輸送の駆動力に静水圧が寄与するか否かを検討した。摘出血管灌流ラット顎下腺を用い、灌流動脈圧を測定しながら、灌流流速を変化させ、水分泌速度、蛍光マーカー(Lucifer Yellow)の分泌を測定した。その結果、1) 灌流流速を変化させ、毛細管床の静水圧を上昇させた場合、蛍光色素分泌速度が静水圧に比例して増加した。2) 高静水圧では水分分泌は比例して増加しなかったが、低圧側の測定では水分泌速度と静水圧は比例し、高圧では水がリークする可能性が示された。即ち、唾液水分分泌のうち大部分を占める傍細胞輸送は、a)灌流圧が駆動力になり溶媒が起動することにより起こる。b)溶媒移動に牽引され水と溶質が移動すると結論された。また、灌流腺を共焦点レーザ顕微鏡ステージにおき、蛍光マーカー(ローダミンデキストラン)を灌流し高速でスライスし、細胞間隙の3次元像を計時的に1秒の時間間隔で撮像した。
ii) 漢方薬の唾液水分分泌増強作用機構:これまで、15種類の漢方薬の唾液分泌増強を起こし3つの分泌パターンに分類され、漢方薬の分類と対応することが確認された。各分類の代表的な漢方薬を用い、エネルギー状態を調べた。漢方薬のみの投与で酸素消費の増加と分泌増強に対応する酸素消費増加が観測された。Na/K ATPaseを阻害するouabainの投与により、増加した酸素消費の一部は減少したことから、一部はNa/K ATPaseの活性化が起こっていることが確認された。
3. エンドソーム-ゴルジ細胞内膜系の生理機能(大橋G)
エンドソーム・ゴルジ細胞内膜系の担う生理機能について研究を行った。
これらの細胞内膜コンパートメント上で機能し、noncanonicalなWntシグナル系を介して平面細胞極性シグナルを制御する分子を同定した。その細胞内膜系での局在を制御する翻訳後修飾を同定し、現在、その機能的意味を解析している。
4. 構造変化型ユニバーサル塩基の開発(片岡G)
相対する塩基が特定の塩基でなくてもDNA二重鎖の形成を維持する核酸塩基、pyrimido[4,5-d]pyrimidine-2,4,5,7-(1H,3H,6H,8H)-tetraone を開発した。この人工の核酸塩基は分子内水素移動による迅速なケト-エノール互変異性化と、プリン型-ピリミジン型塩基構造の配座異性化によって、相対する塩基に呼応して構造変化し、アデニン、グアニン、シトシン、チミンのすべてと塩基対を形成する。概念的に全く新規な核酸塩基であり、様々な生化学ツールとしての利用が期待される。
Copyright (C) 2010 NIPS