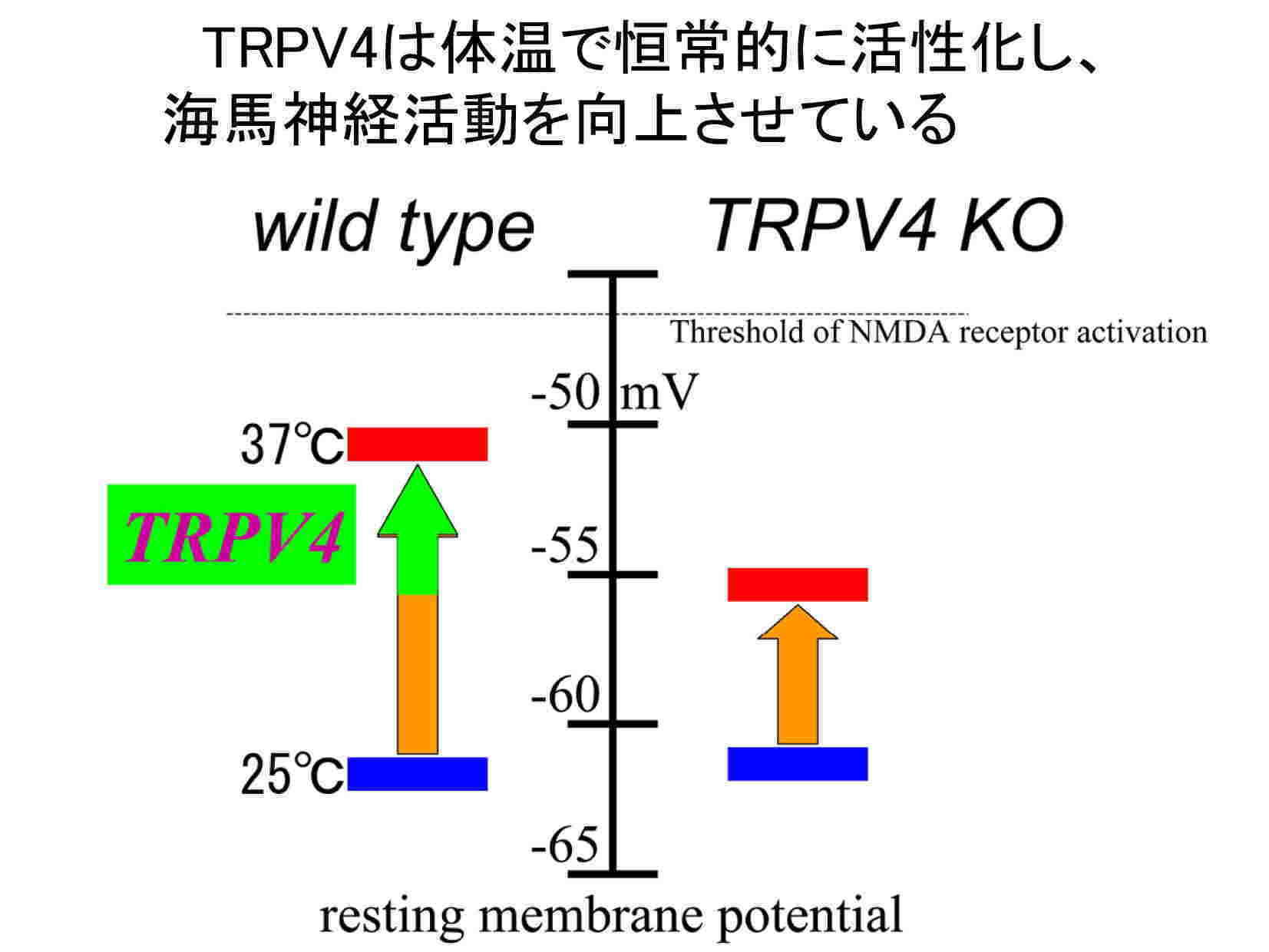
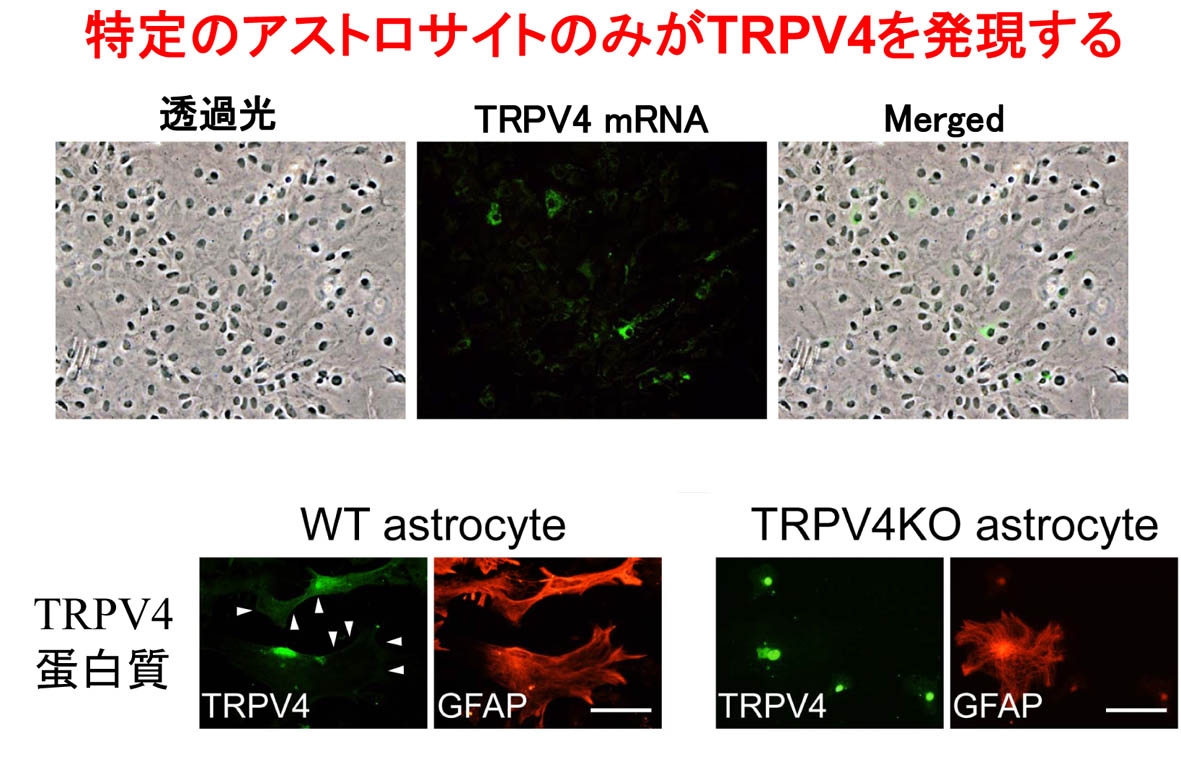
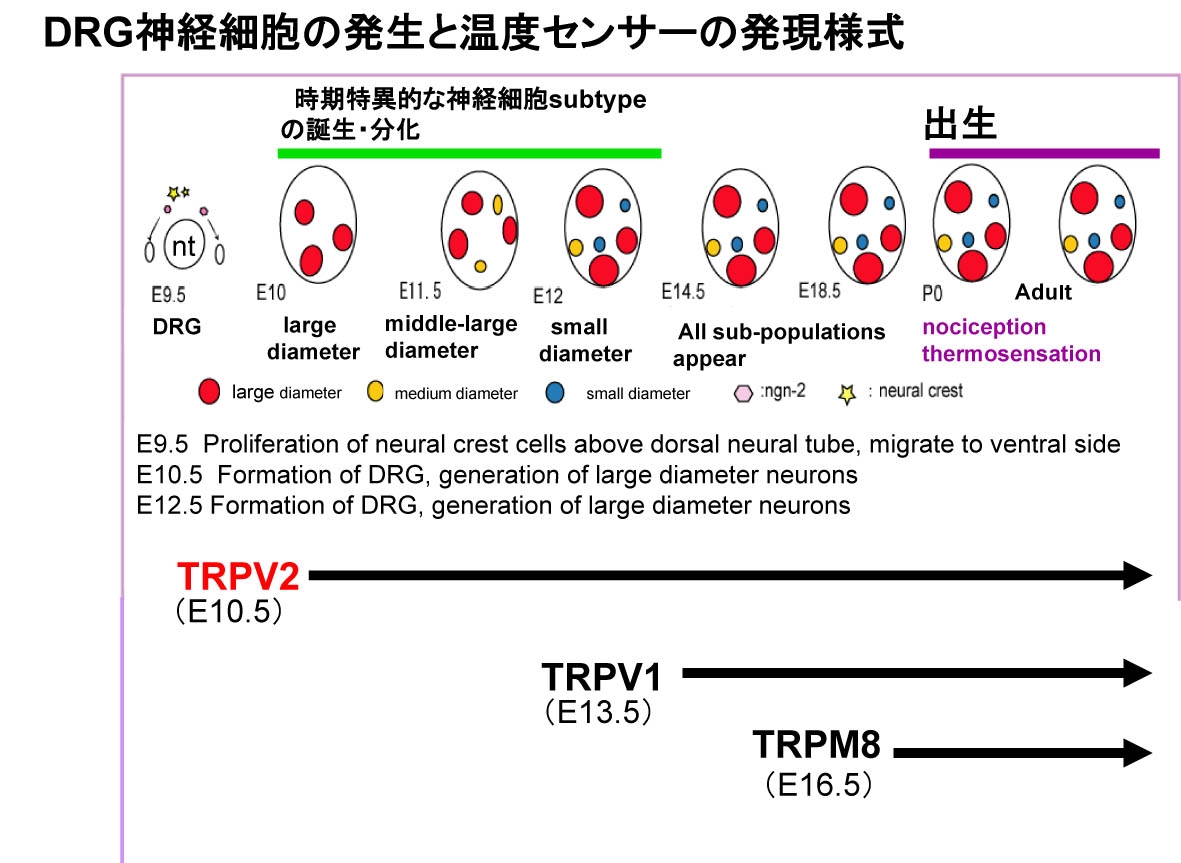
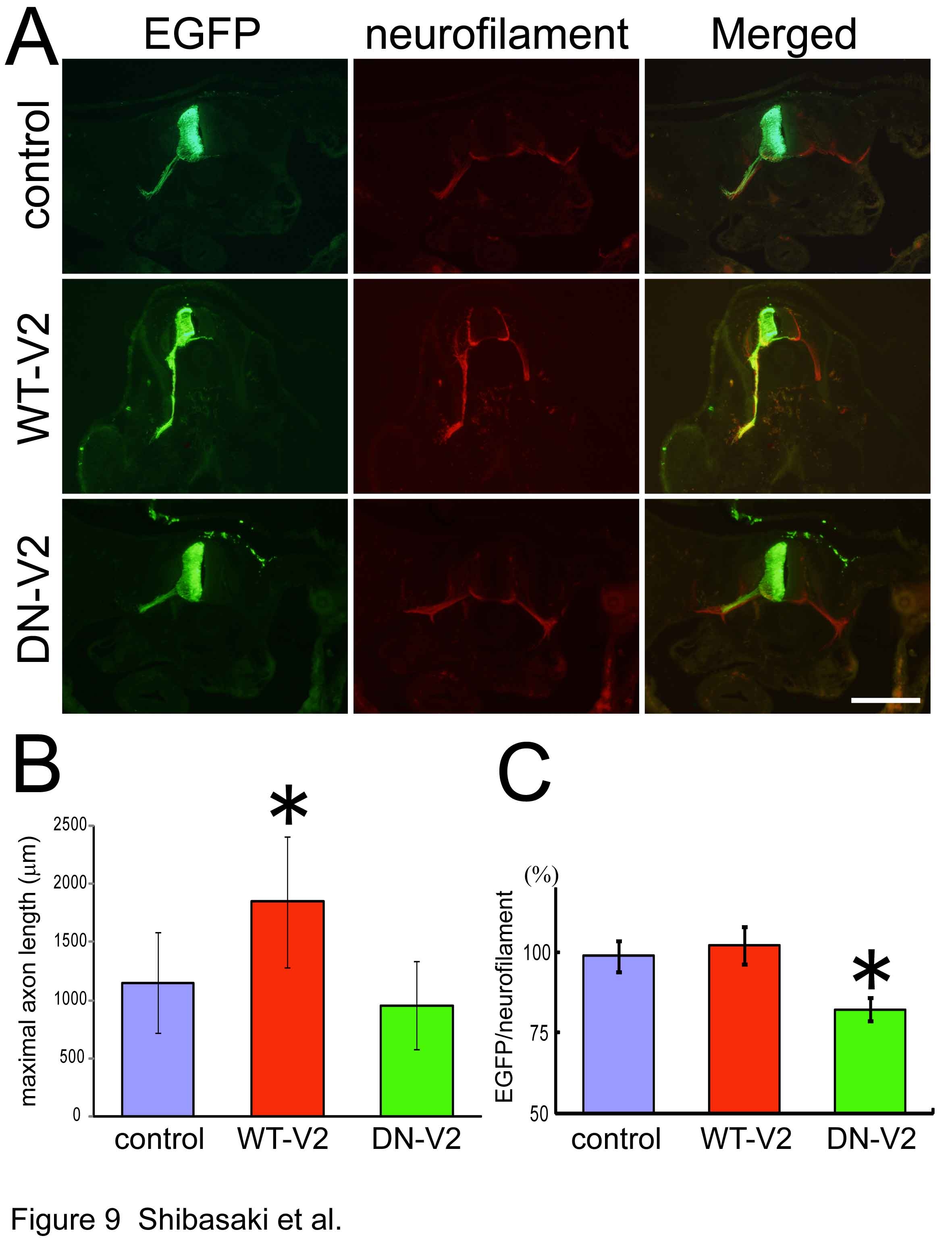
生理学研究所細胞生理部門 HOMEへ
群馬大学大学院医学系研究科脳神経発達統御学講座分子細胞生物学分野 (分子病態学)
共同研究者のHP 京都府立医科大学; 小野勝彦教授 自然科学研究機構 岡崎統合バイオサイエンスセンター 細胞生理部門;富永真琴教授
last up date; 2013/3/14