
研究内容
グリア細胞の発生・分化
中枢神経系を構成する主要細胞であるニューロン、オリゴデンドロサイト、アストロサイトは、いずれも神経上皮細胞と呼ばれる未分化な細胞に由来し ます。この共通の前駆細胞(神経幹細胞)から、まず、ニューロンが生み出され、次いで、グリア細胞が生み出されます。この発生・分化は、さまざまな分泌因 子や転写制御因子により制御されています。私たちの研究室では、いくつかの分化制御分子を独自に同定しており、それらの機能解析を通してグリア細胞の分化 調節メカニズムや生み出された細胞の多様性の形成について研究しています。
主な研究テーマは、以下の通りです。
1)オリゴデンドロサイトの発生とその異常
2)アストロサイトの分化誘導の調節機構について
3)アストロサイトの細胞系譜の解析と機能的多様性
4)神経発生における領域特異化と細胞系譜の解析
それぞれの研究内容を、簡単にご紹介します。
1)オリゴデンドロサイトの発生とその異常
オリゴデンドロサイトの発生は、本研究室の研究テーマの中でもっとも古いものの一つです。池中教授は、ミエリンに局在するタンパクのPLP遺伝子の構造と機能について長年研究しています。鹿川博士(現・熊本大学・助教授)が作製した、PLP過剰発現トランスジェニックマウスは、ミエリンの再生が阻害され、脱髄疾患の有用なモデル動物です。最近では、脊髄背側から分泌されるオリゴデンドロサイト発生を抑制する因子がWntタンパクである事を見いだしました。
小野助教授は、O4抗体がニワトリ胚において非常に初期のオリゴデンドロサイト前駆細胞を染める事を見いだし、視神経内のオリゴデンドロサイトの発生部位、移動経路を同定しました。
竹林助手はオリゴデンドロサイトや運動ニューロンの発生に必須のbHLH型転写因子・Olig1/2を同定し、その機能をOlig2ノックアウトマウス作製により証明しました。
本研究室は、オリゴデンドロサイト研究を行っている研究者が揃っている、日本の中でも希有な研究室であると言えます。
2)アストロサイトの分化誘導の調節機構
レトロウイルスを用いた発現スクリーニングにより、システインプロテアーゼ阻害因子・シスタチンCによって、アストロサイト特異的なGFAPの発現が誘導されることを見出しました。アストロサイトの分化誘導分子の一つであると考えられ、その詳細な機能解析を培養細胞、マウス個体を用いて行っています。
3)アストロサイトの細胞系譜の解析と機能的多様性
ニューロンの多様性は、神経管の背腹軸方向に転写制御因子により形成されるドメイン構造に対応して誘導されることが報告されています。近年、アス トロサイトにもさまざまな機能的な多様性が報告されるようになってきましたが、このアストロサイトの機能的な多様性も、発生期のドメイン構造によって規定 されるのではないかという仮説を立てています。マウス胎仔の脳にエレクトロポレーション法によりレポーター遺伝子を導入して特定の細胞を標識し、その細胞 を成熟するまで追跡してそれらの機能を解析しています。
4)神経発生における領域特異化と細胞系譜の解析
Cre発現プラスミドをエレクトロポレーション法により遺伝子導入する新たな恒久的細胞標識法を開発しました。また、タモキシフェン誘導性CreERノックインマウスを用いた領域特異的・時期特異的な遺伝子組み換え法を用いて、発生期の各ドメイン構造から生み出される細胞系譜について解析しています(図1)。Olig2の機能は、主に脊髄で調べられてきましたが、前脳(図2)や後脳における機能についても調べています。
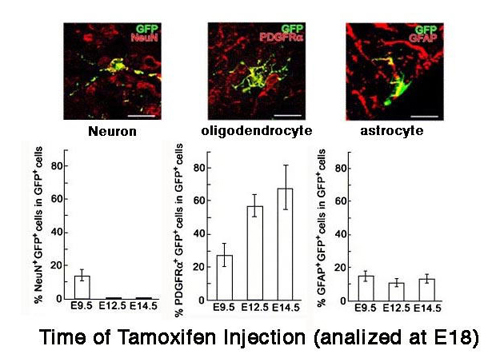
図1
(上段)タモキシフェン誘導性Creを使って、脊髄のOlig2陽性細胞にGFP(緑色蛍光タンパク)を発現させ標識した。細胞種特異的マーカーと の二重染色により、ニューロン(NeuN+)、オリゴデンドロサイト(PDGFRa+)、アストロサイト(GFAP+)に分化する事がわかる。(下段)タ モキシフェンの投与時期を変えることにより、胎生12日以降のOlig2陽性細胞からは、ニューロンではなくオリゴデンドロサイトとアストロサイトが生み 出される事がわかる。
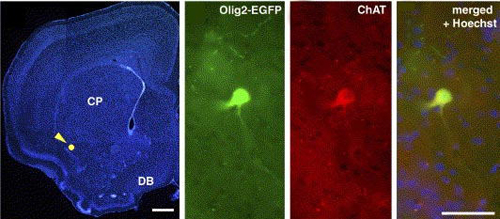
図2
成体マウス前脳の前頭断面(左図;ヘキストで核染色している)。タモキシフェン誘導性Creを使って胎齢12.5日目に前脳のOlig2陽性細胞に GFP(緑色蛍光タンパク)を発現させ標識し、これを成体になった段階で解析したもの。黄色矢頭の部分の細胞を拡大して観察すると、ChAT(コリンアセ チルトランスフェラーゼ)陽性のコリン作動性ニューロンに分化することがわかる(右3枚の図)。