鈴木 誠(自治医科大学薬理学分子薬理学講座)
Sabirov Ravshan
Tweety(TTY)はショウジョウバエの遺伝子で,5 - 6回膜貫通蛋白質である。鈴木らはこの蛋白質がClチャネルをコードしている可能性を示した。即ち,TTY3はChinese Hamster Ovary cellで発現すると,Ca-dependent Cl channelを観察する事ができた。linear I-V, 350 pSでμMレベルのCaを活性に必要とした。Poの電位依存性は脱分極側にあり,非ベル型を示しており,一般的に認められるベル型を示すClチャネルとは 性質を異にしている。TTY1はD,E,Qなどのアミノ酸の繰り返し構造を欠いており,活性化の刺激がわからなかった。しかし,細胞容積を低浸透圧で,大 きくすると活性化した。
TTY1は100-200 pSのチャネルを発現するが,CHO細胞には内因400 pS Clチャネルがあり,測定の障害になった。そこで今回,内因性Cl channelがないHEK-T細胞を用いた。HEK細胞にはTTY3がRT-PCRで検出でき,抗体で蛋白も確認できるが,HEK-T細胞にはTTY 1,2,3共にmRNAは検出できなかった。またHEK細胞には内因性の外向き整流性のClチャネルが見つかるが,HEK-T細胞では見つからなかった。
TTY1の発現をHEK-T細胞で試みたが,発現は認められなかった。そこで,CHO細胞では明らかに発現が確認されたTTY3を用いたが,チャネル発現が認められなかったので,分子自身が膜に移行しているのかどうか検討しなおすことにした。
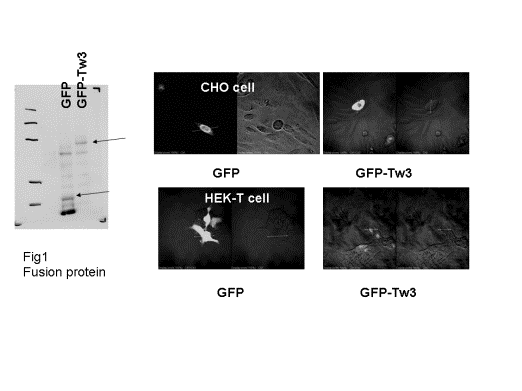
添付図左上にあるようにTTY3をGFP蛋白とfusionさせた。図は,GFPに対する抗体を用い て,ウエスタンを行ったものである。Fusion蛋白ではバンドが上方にシフトしている。これらのベクターを用い,CHO細胞とHEK-T細胞で,発現を 検討した。図右にあるようにGFP単独では蛍光は細胞に一様に広がっている。TTY3-GFPはCHO細胞では膜に発現するのに対し,HEK-T細胞では 細胞質内にとどまっているのが明らかであった。
赤塚結子(三重大学医学部生理学第一講座)
清水貴浩,岡田泰伸
細胞外及び細胞内の浸透圧変化に対応して自らの容積を一定に保とうとする働きは,動物細胞が生命を維持 する上で必要不可欠な機能であるが,最近ではこの容積調節の破綻が細胞死につながることが明らかとなっており,細胞がいかに自らの容積をセンスし対応する かという点に注目が集まっている。細胞が一旦膨張した状態から元の体積に戻る調節性容積減少(regulatory volume decrease: RVD) の過程は,細胞内の蛋白質による情報伝達を介して,最終的には細胞内からのK+とCl-流出が駆動力となって細胞内の水が細胞外に流出することによって達成される。特にこの場合のCl-の通り道であるチャネルは細胞の容積上昇を感知して開口するために容積感受性Cl-チャネル(VSOR)と名づけられているが,最近では正常浸透圧下でアポトーシス誘導剤やH2O2に よってVSORが活性化されることによって,細胞の持続性収縮が起こることが明らかとなり,容積調節だけでなくアポトーシス誘導にも深く関わっていること がわかってきている。VSORの分子実体はいまだ不明であるが,VSOR及びVSORの制御因子はアポトーシスをコントロールするという観点からも重要な 蛋白質であり,これら蛋白質群の分子同定によって細胞の容積調節やアポトーシスのメカニズムについてさらに多くの情報が得られることが期待される。
現在までに報告者らは,VSORの調節蛋白質としてATP-binding cassette (ABC)蛋白質スーパーファミリーに属するABCF2を同定しているが,今回の共同研究によって,ABCF2がVSORの電流を抑制すること と,ABCF2の発現によってRVDの遅延が起こることを見出した。さらに,ABCF2がアクチン結合蛋白質であるアクチニン-4と結合すること,その結 合には両者のアミノ末端側領域が必要であることを明らかにしており,アクチニン-4にはABCF2によるVSORの抑制を解除する働きがあることがわかっ た(投稿準備中)。さらに,ABCF2結合蛋白質の探索によってVSORの候補蛋白質を見出し,その機能解析を進めている。
河田照雄,楠堂達也,永井宏幸,向井佐輝子,浅野亘(京都大学大学院農学研究科)
高橋信之,岡田泰伸
脂肪細胞は「脂肪を貯める」ことが第一義的な生理機能である。そのために容積が約1~2万倍まで肥大化 し,可逆的に維持しうる。このような特性は他の細胞に類を見ない。このことは生体のエネルギー供給の保証ともなり,また動物が獲得した進化特性でもある。 しかしながら,飽食化した現代社会においては,生活習慣病を招来する主要因ともなっている。本研究では,脂肪細胞が,自身でその発達度合いをどのようにモ ニターして,その形態形成・容積維持・肥大化制御を行っているかを分子細胞生物学的に解明することを目的としている。
昨年度までの共同研究において,脂肪滴をため込んだ分化脂肪細胞を使った細胞内カルシウムイメージング の実験系を確立し,脂肪細胞の分化に伴い発現が上昇するTRPチャネルをモニター分子の候補として同定した。同定されたTRPチャネルはこれまでの研究よ り,温度,成長因子だけでなく,ストレッチなども活性化シグナルとすることが報告されており,容積センサーとしての条件を備えていると考えられる。今年度 は昨年度構築したカルシウムイオンイメージング及びパッチクランプでの測定系を用いたTRPチャネルの特性評価および容積センサー分子としての機能の解析 を試みている,まだ具体的な成果は得られていないが引き続き行っていく予定である。また,脂肪細胞が外界の脂肪酸等の化学的因子をシグナルとして受容して いる可能性も考えられる。本チャネルを直接的あるいは間接的に活性化する生理的なリガンドはほとんど同定されていないが,同じサブファミリーに属する TRPチャネルは脂肪酸など様々な化学的因子により活性化されることから,本チャネルも脂肪酸類縁体によって活性化される可能性が高い。そこで,細胞内カ ルシウムイオン濃度に変化を与えるシグナル分子の効率的な探索を行うために,蛍光プレートリーダーを用いたTRPチャネルのリガンド一次スクリーニング系 を確立し現在スクリーニング中である。さらにスクリーニングで得られたリガンド候補物質の確認のため,カルシウムイオンイメージングセット及びパッチクラ ンプセットによる測定法も確立した。今後はこれらの方法を用い,TRPチャネルのリガンド探索を進め,それがTRPチャネルを介して脂肪細胞の容積制御お よび分化に関してどのような役割を果たしているかということも,先の容積センサーとしての解析に加えて行っていく予定である。
檜山武史(自然科学研究機構 基礎生物学研究所)
岡田泰伸,井本敬二
Naxチャンネルは細胞外のNa+濃度の生理的範囲での上昇に応答して開口する Naレベルセンサーである。脳室周囲器官の一つである脳弓下器官において体液Naレベル増加の検出に関与する。本研究では,中枢におけるNaxの発現を詳 細に検討した。また,Naxが細胞生理機能において果たす役割を検討する手がかりの一つとして,細胞内領域に結合する分子を探索した。さらに,Naxの開 口により細胞内へ流入したNaがNa/K-ATPaseの機能制御に関与する可能性,細胞内液の浸透圧変化に伴って細胞容積が変化する可能性について検討 した。
1)Naxの中枢における発現解析
免疫電子顕微鏡法により,Naxが脳室周囲器官の脳弓下器官と終板脈管器官において,上衣細胞と星状細 胞から伸びたニューロン周囲の薄膜状突起に発現することが判明した。脳弓下器官から単離したグリア細胞においてNaイメージングを行なったところ,細胞外 Naレベルの上昇に応じたNa流入が確認された。
2)Naxの細胞内領域と結合する分子の探索
Naxの細胞内領域と相互作用する蛋白質を探索した。成体マウスの後根神経節からcDNAライブラリーを作成し,酵母ツー・ハイブリッド法により結合分子を探索し,Na/K-ATPaseを含む複数の結合候補分子を見出した。
3)Naxによる細胞代謝制御の可能性に関する検討
Na/K-ATPaseとNaxの相互作用の存在が示唆されたことから,グルコース・イメージング法を 用いて脳弓下器官より単離したグリア細胞のグルコース代謝を測定した。Nax陽性細胞においてNa依存的にグルコース取り込みが増加することが明らかと なった。また,マウスの脳から作成した脳弓下器官の急性スライスにおいて,野生型マウス特異的にNa依存的なグルコース取り込みが観察された。
4)Naxを介したイオン流入が細胞容積に影響する可能性についての検討
Naxチャンネルの開口によるイオン流入によってもたらされる細胞内浸透圧の増加が,水分子の流入と細 胞容積の増加をもたらす可能性を検討した。C6グリオーマ細胞にNaxを発現させ,細胞膜をPKH26により染色して共焦点顕微鏡により細胞の形態を観察 した。細胞外液のナトリウム濃度変化に伴って細胞体の大きさが顕著に変化することはなかった。今後,局所的かつ微細な形態変化を捉えられる実験系を構築 し,さらに検討する予定である。
【追加情報】(調査月2007年8月、記入月2008年6月)
関係論文
富永 真琴(岡崎統合バイオサイエンスセンター)
柴崎 貢志(岡崎統合バイオサイエンスセンター)
東 智広(岡崎統合バイオサイエンスセンター)
岡田 泰伸(生理学研究所)
清水 貴浩(生理学研究所)
細胞外の低浸透圧状態を感知してさまざまな細胞応答が引き起こされる。低浸透圧で活性化するクロライド チャネルの電気生理学的な解析はなされているが,いまだその分子実体は明らかでない。一方,低浸透圧刺激によって活性化する非選択性陽イオンチャネル TRPV4がクローニングされ,バイオ分子センサーとして浸透圧感受性に分子から迫ることが可能になった。そこで,まず,TRPV4の低浸透圧感受性を TRPV4を強制発現させた培養細胞で確認した後,TRPV4が発現していることが知られている表皮ケラチノサイトとマウス脳でTRPV4の遺伝子発現を 検討し,十分量のmRNAの発現を観察した。次に,単離ケラチノサイト,単離海馬神経細胞で低浸透圧応答をCa2+イメージング法,パッチクランプ法を用いて検討した。その結果,両方法において,低浸透圧刺激に対する細胞内Ca2+濃 度の増加,膜電流の活性化を観察した。そこで,容積感受性クロライドチャネルがTRPV4と機能的のみならず物理的にカップルしていることを想定して Yeast Two-Hybrid法を用いて,ケラチノサイトおよび脳のcDNAライブラリーからTRPV4のカルボキシル末端と結合する蛋白質を探索した。細胞骨格 関連蛋白質は得られたもの,容積感受性クロライドチャネルの分子実体と思われる蛋白質を得ることはできなかった。TRPV4と結合する胞骨格関連蛋白質と の結合蛋白質として容積感受性クロライドチャネルを得ることができるかもしれない。
森泰生(京都大学大学院工学研究科)
井本敬二(生理学研究所)
小脳プルキンエ細胞においては代謝型グルタミン酸受容体などホスホリパーゼC (PLC)と共役した受容体が,神経可塑性などの生理現象に重要な役割を果たしていることが知られている。しかしこれら受容体活性化の時空間的な動態とそ の制御機構については明らかになっていない。本研究ではPLCの活性化により産生されるイノシトール3リン酸に対する特異的蛍光プローブを用いて,PLC 共役型受容体の活性化をリアルタイムで可視化すること目的とし技術的改良を行った。
これまでに我々は,PLCの活性化により産生されるイノシトール3リン酸特異的蛍光プローブの作製に成功し,その有用性を報告した。本研究では急性単離した小脳プルキンエ細胞に蛍光プローブを取り込ませることを試みた。
PLCのPHに基づいたイノシトール三リン酸特異的蛍光プローブの,現在までの最大の問題点の一つが細 胞内への導入法であった。今回,エレクトロポレーションにより導入効率が数倍上昇することが明らかとなった。また,このことにより様々の細胞に蛍光プロー ブの導入が可能となり,汎用性が格段に改善された。
岡部繁男(東京医科歯科大学・大学院医歯学総合研究科)
培養神経細胞および脳スライスにおいてGFP融合シナプス蛋白を発現させ,これらの蛋白質のシナプス1 個あたりの絶対数をGFPの蛍光量をもとに算出することが可能である。更に内在性の分子についてもGFP融合蛋白質との量比からその単一シナプスあたりの 存在数を推定することが出来れば,機能的な分子数の測定が不可能な多様なシナプス分子についてのその局所に存在する分子数を求めることが出来る。このよう な方法論の具体化を目的として以下の実験を行った。
シナプス後肥厚部とほぼ同じサイズの蛍光ビーズの蛍光量を測定し,全反射顕微鏡による観察から求められ た単一GFP分子の蛍光量と比較を行った。その結果,単一GFP分子と直径200 nmの蛍光ビーズとの蛍光比は1:3700と求められた。このキャリブレーションされた蛍光ビーズを利用して,培養海馬神経細胞の単一シナプスに局在する GFP融合足場蛋白質4種類(PSD-95,GKAP,Shank,Homer) の分子数を算出した。更にGFP融合足場蛋白質を過剰発現する細胞と,していない細胞での足場蛋白質の量比を蛍光抗体法で決定することにより,遺伝子導入 操作を加えていない海馬神経細胞での内在性の分子数を決定した。内在性の分子については,PSD-95,GKAP,Shank,Homerの4種類の足場 蛋白質全てについて,およそ100-450分子が1つのシナプスあたり存在することが明らかになった。4種類の蛋白質の分子数に大きな隔たりが無いことか ら,足場蛋白質間の結合のstoichiometryは比較的単純であることが示唆された。更に培養神経細胞が分化する過程で,単一シナプスあたりの分子 数は培養二週間目までは単調に増加するが,それ以降は平均値およびその分散に大きな変化は見られなかった。一旦成熟した神経細胞においては,単一シナプス での足場蛋白質の分子数およびその分散を一定に保つ分子機構が存在すると考えられた。更にこれら4種類のシナプス足場蛋白質の質量の総和は約100 MDaとなり,PSD構造全体の質量のおよそ10%を占めることになる。この結果は今回測定した4種類の足場蛋白質が形成する骨組がPSD構造の維持に十 分な役割を果たすことを示唆する。
上記のシナプス足場蛋白質の絶対数測定法の確立を基に,この手法を更にシナプスの微細構造および機能と 結び付けるための実験手法の検討を行った。より内在性の蛋白質発現に近い状態での測定を可能にするため,GFP融合シナプス後部蛋白質を発現するトランス ジェニックマウス系統の作製を行い,これらのマウス系統由来の細胞で同様の分子数推定法を適用することが可能であることを確認した。更に海馬神経細胞の培 養に用いる新規基質の検討を行った。新規の基質を利用することで,分子数の推定に必要な蛍光画像を取得後に,電子顕微鏡標本の作製を行い,同一のシナプス を同定することが可能になった。今後この方法を利用して,シナプスに局在する足場蛋白質の絶対数と,スパインおよびPSDの微細構造の関連を解析する予定 である。
【追加情報】(調査月2007年8月、記入月2008年6月)
発表論文
関係論文
永雄総一(理化学研究所脳科学総合研究センター 運動学習制御研究チーム)
慢性マウス標本を用いて,運動学習の短期記憶から長期記憶への固定化の神経機構を実験的に検討した。マ ウスに,1日1時間の視機性眼球反応(HOKR) の訓練を1週間継続的に行うと,その動特性に短期適応と長期適応が生じる。先行研究により,短期と長期の記憶痕跡がそれぞれ小脳片葉と前庭核に形成される ことと,前庭核の長期記憶痕跡の形成に,小脳片葉と小脳神経回路のシナプス可塑性の長期抑圧がともに必要であることが報告されている。本年度は,小脳皮質 の遺伝子発現の変化が,短期記憶と長期記憶の痕跡の形成に関与している可能性を,遺伝子マイクロアレー法を用いて検討した。1日1時間の訓練を行った直後 と,1週間持続的に訓練を行った直後のマウスから小脳皮質を摘出し,片葉とその近傍の傍片葉を分離摘出した。摘出した組織よりmRNAを抽出 し,GeneChip法(Affymetrix)により45,000個以上の遺伝子について,その発現パターンを比較検討した。その結果,長期適応が生じ たマウスの片葉では,対照群のマウスの片葉と適応が生じたマウスの傍片葉に比べて,1,200個程度の遺伝子に発現量の有意な変化が生じており,かつその うちの70%では発現量が減少していることを見出した。一方,短期適応が生じたマウスの片葉では,対照群のマウスの片葉と短期適応が生じたマウスの傍片葉 に比べて,発現量の変化が見られた遺伝子数は400個程度で,そのうちの約3分の2には発現量の減少が見られた。片葉で,短期適応と長期適応にともに平行 して発現の減少する遺伝子群がそれぞれ存在することは,運動学習によって小脳皮質の遺伝子発現にも長期抑圧が生じることを示唆する。これらの結果を 2005年度の冬の日本分子生物学会に報告した。
姜英男(大阪大学大学院歯学研究科高次脳口腔機能学講座口腔生理学教室)
重本隆一(自然科学研究機構生理学研究所大脳皮質機能研究系脳形態解析研究部門)
持続性Na+電流 (persistent Na+ current) は,軸索における興奮性シナプス入力の統合や,三叉神経中脳路核ニューロンを含む様々なニューロンで既に報告がなされている様に,膜電位のオシレーション において重要な役割を担っている。我々は本研究課題において,三叉神経中脳路核ニューロンの軸索小丘から幹軸索にわたる領域において,持続性Na+電流が活動電位の生成及び細胞体への侵入に対して果たす重要な役割を明らかにした。
軸索小丘からホールセル・パッチクランプを行なうと,膜電位のオシレーションが観察されたが,その閾値 は,細胞体において記録されたオシレーションのものよりも10 mV低かった。4-アミノピリジンを投与すると,オシレーションは顕著に遅くなり,5~10 microM リルゾール及び5~10 nM テトロドトキシンに感受性を持つランプ状あるいは持続性の脱分極が観察された。細胞体に0.5 mM QX-314を含んだ内液を充填したパッチ電極を,軸索小丘にはQX-314を含まない内液を充填したパッチ電極を用いてデュアル・ホールセル・パッチク ランプを形成することにより,細胞体から軸索小丘にかけてQX-314の濃度勾配が生じると想定され,この時,短い電流パルスを繰り返し与えると,電流パ ルスによって生じる遅い閾値下脱分極の持続時間のみならず,軸索小丘で記録される活動電位の振幅も,時間の経過とともに減少していった。一方,細胞体で記 録される活動電位はほとんど変化しなかった。これらの所見から,持続性Na+電流は,軸索小丘部或いは,それを越えた幹軸索において生成されている可能性が示された。更に,軸索小丘部のパッチ電極を通じた電流パルス注入,及び,幹軸索の電気刺激を交互に繰り返して活動電位を発生させると,10~50 nM テトロドトキシン或いは10 microM リルゾールの灌流投与によって,先ず幹軸索刺激による軸索活動電位の発生が抑制される。その際,電流パルスの注入によって活動電位を生成することはできるが,必要な電流強度は段階的に上昇していた。その後は,活動電位の逆伝播 (spike backpropagation) が観察されなかった。これらの所見から,持続性Na+電流は,軸索小丘から幹軸索にわたる領域に局在し,活動電位の生成及び侵入に直接的に関与していることが示唆された。
【追加情報】(調査月2007年8月、記入月2008年6月)
発表論文
関係論文
福田 敦夫(浜松医科大学)
鍋倉 淳一(生理学研究所)
KCC2は細胞内のCl-を細胞外に排出することにより[Cl-]iを低く維持して,GABAの作用を過分極すなわち抑制性ならしめる蛋白であるが,細胞内でのKCC2の機能調節については今だ不明な点が多い。yeast two-hybrid法,免疫沈降法を用いて,脳型のクレアチンキナーゼ(CKB)をKCC2機能制御タンパク候補として同定した。機能的相互作用を見い出すため,HEK293細胞にKCC2とCl-チャンネルであるグリシン受容体を共発現させたKCC2活性の評価系を用いて,KCC2の活性の指標である[Cl-]iを推定できるグリシンの逆転電位(Egly)をグラミシジン穿孔パッチクランプ法を用いて測定した。この系にdominant-negative CKBを導入するとKCC2発現細胞においてのみEglyは脱分極側にシフトし,wild type導入細胞より有意に[Cl-]iが上昇した。また,ネイティブにKCC2とCKBが発現しているマウス大脳皮質の初代培養細胞を用いてCKBの抑制実験を行った。CKB阻害剤のDNFBを投与するとり[Cl-]iが有意に上昇した。これらのことから,CKBはKCC2を活性化することが示唆された。
回路再編でのCl-ホメオスタシスの役割をバイオCl-セ ンサーとしてのGABA/グリシン応答を指標に解析する目的で,まず回路形成過程でのGABA/グリシン応答の役割を検討した。大脳皮質層構造構築に重要 な役割をもつ辺縁帯の細胞間のクロストークにおけるGABA/グリシン応答の役割を明らかにするため,辺縁帯における興奮の空間的伝播を膜電位イメージン グで可視化した。活動電位は径シナプス的に放射状に伝播し,GABAA受容体とグリシン受容体が関与していたがグルタミン酸受容体は関与していなかった。細胞内にCl-を取り込み[Cl-]iを 高く維持してGABAの作用を脱分極すなわち興奮性ならしめる蛋白NKCC1の活性を阻害すると興奮伝播は抑制された。マイクロダイアライシス法を用いて アミノ酸を測定したところ,単発刺激で内因性のグリシン受容体アゴニストであるタウリンとGABAが放出されていた。すなわち,NKCC1によって高 [Cl-]iを維持する辺縁帯の細胞では,タウリンとGABAが興奮性に働いて細胞間クロストークに寄与していた。
【追加情報】(調査月2007年8月、記入月2008年6月)
発表論文
関係論文
岡田 誠剛(関西医科大学)
鍋倉 淳一(生理学研究所)
神経活動は発達段階におけるシナプス形成・排除に影響を与え,周囲の神経細胞との活動性の差の重要性が 近年の研究によって示されている。一方,神経活動の同期性は,中枢シナプス伝達の可塑的変化や,視床・大脳皮質間の情報伝達に対して重要な役割を果たすこ とが示されている。これまでの可塑性についての研究は,NMDA受容体が同期性センサーとして作用することを示しているが,同受容体の発現量は部位,時期 によって大きく異なり,これ以外の分子も同期性センサーとして関与している可能性が推測される。内向き整流性K+チャンネル (Kir)mRNAの発現は中枢神経系において広く認められるが,その作用は明らかではなく,Kirの内向き整流性およびシナプス後膜に存在する PSD95への結合能から,同チャンネルがシナプス伝達に対して同期性センサーとしてはたらいている事が推測される。そこで,本研究は内在性Kir発現が 少ない(Prüss et al,2005)海馬CA1領域の錐体細胞に,低毒性で長期間の発現が可能なレンチウイルスベクターを用いて,Kirを発現させ,神経活動への影響を全細 胞記録により検討した。Kir電流は,未感染細胞では3.9+1.0 nSであったのに対し,ウイルスベクター感染細胞では27.1+4.1 nSであった。静止膜電位はKir発現により,平均8.8 mV低下した。mEPSPによる電位上昇は,Kir発現により有意に低下した。さらに,電流注入による電位変化を検討すると,200pAまでは,Kir発 現は上昇を有意に抑制したにもかかわらず,300 pA以上の電流注入では抑制は認められなかった。すなわち,Kirの内向き整流性が,興奮性入力に対して同期性センサーあるいはノイズ・フィルターとして の作用することが示唆された。また,予備的な実験ではKir発現は,幼弱な海馬に特徴的に認められるGiant Depolarizing Potentialsの頻度を減少させた。本研究により,Kirの同期性センサーとしての作用が示されたと共に,同発現ベクターによる神経活動の抑制作用 は,神経活動およびその同期性が,神経系の発達や高次機能へ与える影響の今後の検討に有用である事が示唆された。
吉村 恵,古江 秀昌(九州大学)
鍋倉淳一(生理学研究所)
唐辛子の主成分であるカプサイシン受容体(TRPV1) が遺伝子クローニングされ,機能的発現実験からカプサイシンのみでなく,熱およびプロトンの変化を受容することが明らかになった。この受容体は神経末梢の みでなく脊髄内中枢端にも発現しており,何らかの役割を果たしていることが示唆されていた。そこで,脊髄スライスに後根を付した標本とin vivoの標本からパッチクランプ記録を行い,カプサイシンに対する作用,pH 変化に対する応答およびin vivo標本を用い皮膚に熱刺激を加えた時の応答の解析を行うことを目的にした。スライスパッチクランプ記録:成熟ラット脊髄の横断スライスに後根を付し た標本を用い,後角第II層の膠様質細胞からパッチクランプ記録を行い,後根刺激によって誘起されるシナプス応答を対照に,カプサイシン,pHおよび熱変 化に対する応答を記録解析した。
In vivoパッチクランプ記録:ラット腰部脊髄の椎弓切除を行い,脳脊髄固定装置にセットする。脊髄表面をKrebs液で灌流し,薬液も同じラインから投与 を行った。パッチ電極を膠様質に刺入し,皮膚刺激によって誘起されるシナプス応答を記録解析した。記録側の後肢に触および機械的痛み刺激を加える と,EPSCの振幅および頻度の著明な増大が全ての細胞で観察された。しかしながら,熱刺激を加えても全ての細胞で何ら応答を得ることが出来なかった。と ころが,カプサイシンを脊髄に直接投与するとほとんど全ての細胞でmEPSCの頻度の増加が観察され,膠様質細胞に入力する線維の中枢側にはTRPV1受 容体が発現されていることが示唆された。一般的な考えでは中枢側に発現している受容体は同じ線維の末梢側にも発現している,すなわち熱刺激にも応答するこ とが予測されたが,結果は一般的な予測とは矛盾するものであった。さらに熱刺激を加えることによって膠様質細胞にC-Fos の発現が増える報告があり,そのこととも矛盾している。このことはC-Fosの発現には必ずしもシナプス伝達が必須ではないことを示唆している。次に,熱 刺激情報が如何なる部位に運ばれているかを検討するため,深層の細胞から記録を行った。約20%のIII-IV層の細胞では熱刺激によって著明なEPSC の頻度の増大が観察された。この応答は全て経過の早いEPSC,すなわちグルタミン酸によるものであり,ペプタイドの関与はないものと判断された。
【追加情報】(調査月2007年8月、記入月2008年6月)
関係論文
石橋 仁(九州大学)
張 一成,鍋倉淳一(生理学研究所)
活動電位がシナプス前神経終末部に到達すると,電位依存性Ca2+ チャネルが開口し,細胞外からCa2+ が流入して神経伝達物質が放出される。従って,細胞外Ca2+ 濃度が低下すると神経伝達物質の放出は減少すると考えられている。一方,シナプス周囲の細胞外スペースは非常に狭いため,シナプス伝達に伴うシナプス前終末部およびシナプス後細胞の細胞内へのCa2+ 流入によって,細胞外Ca2+ 濃度は急速に減少すると考えられている。しかし,これまで,細胞外のCa2+ 濃度が低下した場合に神経終末部が受ける影響は十分には解明されていなかった。
我々は,細胞外にCa2+ が存在しなくても高K+ 溶液による脱分極刺激によって神経伝達物質の放出が増強されることを最近発見し,予備実験の結果からホスホリパーゼC (PLC) の関与が示唆されていた。本年度は,この現象の詳細なメカニズムを解明することを目的に研究を遂行した。ラット脊髄後角からシナプス前神経終末部が付着し た状態で急性単離した神経細胞に,ホールセルパッチクランプ法を適用してグリシン作動性の自発性抑制性シナプス後電流(IPSC)を記録し,細胞外のK+ 濃度をコントロールの2.5mMから30mMにするとIPSCの発生頻度が著明に増加した。この応答は,膜透過性Ca2+ キレート剤BAPTA-AM,Ca2+ ポンプ阻害剤Thapsigargin および PLC阻害剤U-73122によって抑制された。U-73122の不活性化体である U-73343は無効であった。従って,細胞外にCa2+ が存在しなくても,神経終末部の脱分極自体によって,細胞内Ca2+ 貯蔵部位からのCa2+ 放出が起こって神経伝達物質の放出が増強されることが明らかとなった。PLCはIP3の濃度上昇を介してCa2+ 放出を誘発することが知られているが,膜透過性IP3受容体拮抗薬と報告されているXestospongin Cおよび2-APBは無効で,Ca2+ 放出の機序は未解明のまま残った。今後,PLC活性化やCa2+ 放出の詳細なメカニズムを明らかにするとともに,生後発達による影響を検討することを予定している。
門脇 孝 (東京大学大学院医学研究科糖尿病代謝内科)
窪田 直人(東京大学大学院医学研究科糖尿病代謝内科)
窪田 哲也(東京大学大学院医学研究科糖尿病代謝内科)
岡本 昌之(東京大学大学院医学研究科糖尿病代謝内科)
矢野 互 (東京大学大学院医学研究科糖尿病代謝内科)
箕越 靖彦
アディポネクチンは,レプチンとともに脂肪細胞から分泌されるアディポカインである。レプチンは中枢で は生体内エネルギーセンサーである視床下部AMPK活性を低下させることにより摂食を抑制し,末梢では骨格筋におけるAMPK活性を上昇させ脂肪酸酸化を 促進させる。一方アディポネクチンも肝臓,骨格筋においてAMPK活性を上昇させ脂肪酸酸化を促進し,インスリン感受性ホルモンとして作用することが明ら かとなっているが,中枢における役割はまだ不明である。
我々は,まずはじめに,視床下部におけるアディポネクチン受容体の発現について検討した。2つのアディ ポネクチン受容体,AdipoR1/AdipoR 2はいずれも視床下部においてその発現が認められ,その程度は肝臓におけるAdipoR1/AdipoR 2の発現量に匹敵する程であった。さらにその局在をin situ hybridizationにて検討したところ,AdipoR1/AdipoR 2の発現はいずれもレプチン受容体が強発現している視床下部の特に弓状核に強く認められた。次に我々は,髄液中にアディポネクチンが存在するかどうかにつ いて検討した。同じ個体の血清と髄液からそれぞれサンプルを採取しその濃度を測定したところ,髄液中のアディポネクチン濃度は血中の約1/2000程度存 在することが明らかとなった。さらに摂食とアディポネクチンの関連について検討したところ,血中のレプチン濃度とは逆に,血中のアディポネクチン濃度と視 床下部におけるAdipoR 1の発現は絶食時に上昇し,逆に摂食後低下することが明らかとなった。一方AdipoR 2の発現は絶食,摂食後で有意な変化は認められなかった。以上のことより,アディポネクチンは中枢,特に視床下部において摂食さらには個体のエネルギー調 節に重要な役割を果たしている可能性が示唆された。
現在これらの結果を踏まえ,アディポネクチンの中枢作用をより詳細に検討するため,外来性にアディポネ クチンを脳室内に投与し,摂食量やエネルギー代謝量,体重などにおける影響を検討中である。またアディポネクチンによって視床下部AMPK活性が変化する かどうか,もし変化が認められた場合にはレプチン,アディポネクチンの視床下部AMPK活性調節における相互作用についても検討していく方針である。
益崎 裕章(京都大学大学院医学研究科内科学講座内分泌代謝内科)
中所 英樹(京都大学大学院医学研究科内科学講座内分泌代謝内科)
田中 智洋(京都大学大学院医学研究科内科学講座内分泌代謝内科)
石井 崇子(京都大学大学院医学研究科内科学講座内分泌代謝内科)
泰江 慎太郎(京都大学大学院医学研究科内科学講座内分泌代謝内科)
箕越 靖彦
肥満者の多くにおいては,レプチンのエネルギー代謝亢進作用が充分発揮されずレプチン抵抗性の存在が示 唆される。本研究で我々は,レプチン感受性と骨格筋AMPキナーゼ(AMPK)の関連を解明する目的で,高レプチン血症を呈するレプチン過剰発現トランス ジェニックマウス(LepTg)を用い,レプチン感受性の異なる条件下での代謝パラメーターと骨格筋AMPK活性を検討した。レプチンによる体脂肪量の減 少,インスリン感受性の亢進を認め,レプチン感受性を示す標準食下LepTgのヒラメ筋では,対照群と比べてAMP/ATP比の上昇(1.6倍),リン酸 化AMPKの増加(1.5倍),AMPKの標的分子:アセチルCoAカルボキシラーゼ(ACC) のリン酸化の亢進(2.7倍)が認められ,AMPKの持続的活性化が証明された。LepTgは4週間の高脂肪食負荷により,対照群と同程度の肥満とインス リン抵抗性を来したことからレプチン抵抗性の発症が示唆され,この時LepTgのヒラメ筋におけるAMPK活性亢進は消失した。更にLepTgを標準食に 戻すと,対照群と比較し,より急速な体重減少とインスリン感受性亢進状態の回復を示し,レプチン感受性の回復と共に,ヒラメ筋AMPK活性の亢進が認めら れた (Tanaka T et al. Diabetes, 54: 2365, 2005)。以上より,レプチン作用と骨格筋AMPK活性の密接な関連が示され,レプチン感受性の指標としての骨格筋AMPK活性の意義が明らかとなっ た。
次に我々は,長鎖脂肪酸の中枢投与が視床下部のStat3のリン酸化を惹起し摂食を抑制することを明ら かにした。脂肪酸によるStat3のリン酸化はレプチン受容体欠損db/dbマウスや内因性のメラノコルチン受容体拮抗物質:agouti蛋白を視床下部 に異所性に発現するKKAyマウスにおいても認められることから,レプチン受容体シグナル系とは独立した経路を介していると考えられる(投稿準備中)。最近,短鎖脂肪酸であるaリ ポ酸がレプチンと同様に視床下部AMPK活性の抑制を介して摂食を抑制することが報告され,脂肪酸によるエネルギー代謝制御におけるAMPKの意義が注目 される。我々は現在,長鎖脂肪酸-G蛋白共役型受容体システムによる食欲調節機構の解析を進めており,この経路におけるAMPKの生理的意義の解明が期待 される。
【追加情報】(調査月2007年8月、記入月2008年6月)
発表論文
中里 雅光(宮崎大学医学部内科学講座神経呼吸内分泌代謝学分野)
グレリンは,胃で産生される消化管ペプチドで,末梢投与により摂食亢進に作用する。末梢産生物質が摂食 調節に作用するためには,何らかの伝達経路を介し,その情報が中枢に到達する必要がある。我々は,グレリンの摂食亢進作用が,迷走神経遮断および中脳切断 ラットにおいてキャンセルされること,末梢投与したグレリンは,迷走神経求心線維に存在するグレリン受容体に結合し,その電気活動を変化させることにより 空腹情報を視床下部へと伝達することを証明している。さらに,消化管ペプチド;peptide YY (PYY)の摂食抑制機構およびグレリンとcholecystokinin (CCK)の相互作用についての研究から,エネルギーバランス制御における迷走神経求心路の重要性を呈示してきた。グレリン,PYY,CCKの摂食調節に 関するシグナルは,迷走神経経由で延髄孤束核に到達し,ニューロンを変換して視床下部へ伝達されると考えられる。延髄孤束核は,迷走神経をメディエーター とする末梢の物理・化学的刺激あるいはホルモンなどの液性因子情報の入力部位であり,視床下部に投射する多数のノルアドレナリン(NA)産生ニューロンを 含んでいる。我々は,グレリン末梢投与による孤束核でのNA生合成および視床下部弓状核でのNA分泌を評価し,神経解剖学的知見と併せて,グレリンによる 摂食行動とNA神経系との機能連関を検討した。
グレリン投与により視床下部弓状核でのノルアドレナリン放出が増加し,延髄孤束核でのDBH遺伝子発現も有意に増加することを見いだした。弓状核でのNAレベルの上昇は,中脳切断ラットでは認められなかった。a 1およびb 2アドレナリン受容体拮抗薬の前投与により,グレリン投与による摂食亢進作用は減弱した。視床下部弓状核に抗DBH抗体をconjugateした saporin神経毒(DSAP)をマイクロインジェクションし,弓状核でのNAを選択的に枯渇させたラット(DSAPラット)では,孤束核NAニューロ ンの約60%が脱落しており,グレリンによる摂食亢進作用はキャンセルされた。グレリンはNPYニューロンを活性化し,その約50%がDBHの投射を受け ていた。
以上の実験結果から,延髄孤束核に入力した末梢グレリンシグナルは,NA神経系に変換され,弓状核 NPYニューロンを活性化することにより摂食亢進に機能することが明らかになった。胃内分泌細胞から分泌されるグレリンの視床下部への一次,二次神経を介 する情報伝達経路の全貌が初めて明らかになった。
鈴木 敦詞,安田 啓子,関口 佐保子,小野 保長(藤田保健衛生大学・内分泌代謝内科)
長尾 静子(藤田保健衛生大学・疾患モデル教育研究センター)
平林 真澄,加藤 めぐみ(生理学研究所・遺伝子改変動物作製室)
老化に伴う変化では骨組織より軟部組織へのカルシウムの移行がその一因と想定されている。石灰化の担い 手はカルシウムとリンであるが,最近細胞膜Ⅲ型Na依存性リン酸輸送担体Pit-1 の骨ならびに血管石灰化での重要性が注目されている。本研究で作成したPit-1過剰発現ラット(Pit/Tg) は,骨格の成長には影響を認めないものの,二重X線吸収法 (DEXA) 法による骨塩定量にては,雌雄ともに野生型 (WT) に比し骨密度の低下を認めた。また,生後8週目頃より巣状の糸球体障害が明らかとなり,低アルブミン血症,蛋白尿,高脂血症というネフローゼ症候群パター ンをしめし,低栄養状態となって生後8ヶ月で死亡した。末期には低アルブミン血症に基づくと思われる骨軟化症へと進行するが,骨量の低下は低アルブミン血 症発症前から認められることより,Pit-1の過剰発現による細胞へのリン負荷は,骨石灰化に負に働くことが示唆された。また,今回のラットでは,過剰発 現させる細胞を条件付けせずにユビキタスに過剰発現を行ったため,リンの再吸収に関わる腎近位尿細管中でも発現が上昇することが考えられたが,免疫組織学 的検討でも,尿細管細胞でのPit-1蛋白の発現上昇が確認された。そのため尿細管でのPit-1過剰発現により,血中のカルシウム・リン代謝に影響が有 るかどうかについても検討した。末期まで血清カルシウム・クレアチニンの上昇は認めないままであったが,血清リン濃度は常にPit/TgでWTに比べ高値 を示し,体液中のリン濃度の恒常的な上昇により細胞へのリン負荷が持続することが示唆された。
細胞外からの細胞へのリン供給はATP産生のために必須であるが,同時に細胞外リン濃度の上昇による過 剰なリン負荷はアポトーシスを引き起こすことが知られている。今後は,Pit-1過剰発現による細胞障害のメカニズムを解明するために,骨芽細胞ならびに 腎糸球体上皮細胞の初代培養細胞を用いた検討を行う予定である。
八木健,平林敬浩,金子涼輔(大阪大学大学院・生命機能研究科)
平林真澄(生理学研究所・遺伝子改変動物作製室)
CNR (Cadherin-related neuronal receptor) は,非受容体型チロシンリン酸化酵素Fynとの結合活性によって単離された新規カドヘリン様細胞接着分子である。このCNRには14種のファミリー分子が 存在し,いずれも中枢神経系で発現している。CNR遺伝子は染色体上にタンデムに並んだ14個の可変領域エクソンと3つのエクソンからなる共通領域からな るクラスター構造を有し,各CNRファミリーはそれぞれひとつの可変領域エクソンと3つのエクソンからなる共通領域エクソンから転写されていることが明ら かになった。この転写様式はT細胞受容体やイムノグロブリン遺伝子群と類似していることから,CNRはシナプスでの選択的細胞接着と多様化機構の両特徴を 兼ね備えた分子であり,中枢神経系における多様化と組織化をもたらす分子である可能性が考えられる。また,単一神経細胞におけるCNR各分子種の発現様式 をRT-PCRを用いて染色体レベルで解析結果,一つの神経細胞では CNRは複数の分子種を発現しており,その組み合わせは個々の神経細胞ごとに異なっていた。また,各分子種について染色体由来を調べると,片方の染色体の みに由来するものが多数であった。これらは中枢神経系における新たな染色体遺伝子発現機構であり,神経細胞の多様性に寄与していると考えられる。そこで本 研究ではCNRの特異な転写制御機構を明らかにするために種々のCNR遺伝子トランスジェニック (Tg)マウスの作製を行った。なお,CNR遺伝子は全長が200kb以上におよぶため,本研究でのTgマウスを作製は同遺伝子領域を含むBACを改変し たものを導入遺伝子として用いた。
まず,CNR各分子種のエクソンに蛍光タンパク質の遺伝子を挿入し,CNRとの融合タンパク質として発現するように改変したBACを作製し,これを用いてTgマウスを作製した。また,ヒトおよびマウスのゲノム配列を比較したところ,CNR遺伝子の上流に約5 kbpにわたる領域が高度に保存されていることが明らかになった。この領域はCNR 遺伝子の発現制御に関わっていることが予想されるために,この領域を欠損したBACを作製し同様にTgマウスを作製した。
いずれのTgマウスも複数の系統が得られており,現在これらを用いて蛍光を指標にしたCNRの発現様式の解析を進めている。
【追加情報】(調査月2007年8月、記入月2008年6月)
関係論文
片上 秀喜(宮崎大学医学部第3内科)
平林 真澄,加藤 めぐみ(生理学研究所遺伝子改変動物作製室)
成長ホルモン(GH)の作用は肝臓で作られるソマトメジンC (IGF-1)を介するものとされている。血中IGF-I濃度非依存性のGHの各臓器への直接作用は明らかではない。遺伝性侏儒症ラット(dr) は本邦で発見されたGH単独完全欠損症のモデル動物で,天然に存在するGHノックアウトラットである。本研究では脂肪細胞の発生・分化と機能にあたえる GHの組織特異的影響を明らかにするため,自然界に存在するGH・IGF-Iノックアウトラットであるdrに着目し,ヒトGH遺伝子を脂肪細胞に特異的に 発現させ,その生物作用を検討した。ヒトleptin遺伝子上流域-3.6kbpとヒトGH遺伝子2.1kbpのキメラ遺伝子 (Lep-hGH;7.6kbp) を調製し,drラット精子と混合して未受精卵子に顕微注入することにより3匹のLep-hGH-dr個体を作製した。
平成17年度は,成熟個体に達した♂ラットよりとdr♀とを交配し,F1,F2世代を作製し,遺伝子発現と分泌動態を検討した(N=6)。RT-PCR 解析の結果,Lep-hGH遺伝子は脂肪細胞とそれ以外に,♂では精巣,♀では卵巣に強く発現した。生後6ヶ月では♂♀とも,体重は対照drと比較して3-5倍を示し,体脂肪は同週齢のSD♂14-18%に対して,dr♂40-50%,Lep-hGH-dr 68-75%と著しい肥満を示した。20分毎8時間の血中hGH濃度は,対照♂ラットでは2.5~3.5時間毎の規則正しい脈動的分泌 (0.3-100ng/ml)を示したが,Lep-hGH-drでは脈動的分泌は消失し,著しい持続的低値(0.03ng/ml)を示した。
以上の成績より,Lep-hGH-drは予想に反して肥満症の表現型を示した。肥満の機序として,半定 量的RT-PCR解析の結果,脂肪細胞におけるhGH発現量が精巣や卵巣組織の1/5と低値を示し,且つ,ごく低濃度の血中GHが摂食中枢を刺激し,肥満 症を生じた可能性が考えられる。今後は,脂肪細胞のみに強発現するpromoter遺伝子を選択し,再度Tg作出を試み,GHのIGF-1非依存性の脂肪 分解や脂肪細胞分化に及ぼす影響を検討する必要があるものと結論した。
岡 良隆(東京大学大学院理学系研究科)
阿部 秀樹(東京大学大学院理学系研究科)
赤染 康久(東京大学大学院理学系研究科)
羽田 幸祐(東京大学大学院理学系研究科博士課程)
岡村 康司
本計画の提案者らは,ペプチドGnRHを産生するニューロン群の示す神経修飾のメカニズムについて多角的に解析し,多くの業績を上げてきた。特に,GnRHニューロンの示すペースメーカー活動は神経修飾作用の基礎となっていると考えられ,これには新規Na+チャネルの寄与がわかっている。一方,魚類の視床に存在する神経核のニューロンでは特殊な新規Na+チャ ネルが神経核構成ニューロンの応答特性を決定していることがわかっている。しかしながら,それらのチャネルの構造や機能の実態は明らかでない。そこで,統 合バイオサイエンスセンターと東大が連携し,この新規イオンチャネルの分子的実態や生理機能について解析すると同時に行動の動機付けへの関与についても調 べることを計画した。従来の研究で用いてきた熱帯魚ドワーフグーラミーの脳においてNa+チャネル遺伝子のクローニングを行った結果,TTX耐性のNa+チャネル遺伝子断片と思われるものが見出された。今後はこれがGnRHニューロンの示すペースメーカー活動に関与しているかどうかをHeterologous expression実験などで確認すると同時に配列情報を詳細に調べていく予定である。
【追加情報】(調査月2007年8月、記入月2008年6月)
関係論文
吉田 学(東京大学理学系研究科大学院)
柴小菊(東京大学理学系研究科大学院)
稲葉一男(筑波大学下田臨海実験センター)
保住暁子(筑波大学大学院生命環境科学研究科博士課程)
西野敦雄(日本学術振興会特別研究員)
佐藤裕公(筑波大学大学院生命環境科学研究科博士課程)
紺野 在(筑波大学大学院生命環境科学研究科修士課程)
海津麻衣子(筑波大学大学院生命環境科学研究科修士課程)
岡村康司
東大の吉田らは,これまで尾索動物カタユウレイボヤにおいて卵放出性の精子誘引物質SAAFを同定する など,精子運動機能の発現やその制御に関する研究を推進してきた。一方,村田らが同定したCi-VSPはカタユウレイボヤの精子に発現していることから, 精子の運動や受精反応にこの分子が関わる可能性が考えられた。本研究の最終目標は,生殖機能,精子生理機能に関して優れた実験系であるカタユウレイボヤを 用いて,Ci-VSPの生理機能を解明することである。この解析のためには,カタユウレイボヤ精子の鞭毛運動パターンを数理的に解析できる実験系を立ち上 げることが必須である。そこで,ストロボ画像取得システムを用いて精子の運動の軌跡を解析することを行った。
まずは鞭毛運動解析装置の構築を行った。ストロボ装置として高速度LEDを用い,高速度ビデオカメラと 同期を行うことで,200フレーム/秒のレートで高解像度の画像を得ることに成功し,さらに自作の画像解析プログラムを開発することにより,運動中の精子 の鞭毛運動を数理的に解析する手法を確立した。この手法を用い,SAAFが正常精子の鞭毛運動に与える影響の解析を行った。カタユウレイボヤ精子は通常は 円運動をしているが,卵やSAAFの様な誘引源があると,誘引源に近づいている時には直進に近い軌跡を描き,遠ざかっている時には誘引源へ向かうように 「turn」と呼ばれる急激な方向転換を起こすという運動変化を見せる。この際の鞭毛運動パターンの解析を行ったところ,turnの際に一過的にきわめて 非対称的な鞭毛打を起こし,その後対称的なパターンを示すことが明らかとなった。一方,運動中の精子に均一濃度となるようにSAAFを添加すると,円運動 を続けるものの,その運動軌跡の半径が増大していることが明らかとなった。この際,鞭毛運動パターンはより対称的になっていた。この結果は,精子走化性時 に特徴的に見られるturnはSAAFの絶対濃度でコントロールされるのではなく,濃度変化によって引き起こされることを示唆する。
今後,Ci-VSPの変異体を発現させた精子を用いて,この解析法を用い,精子運動におけるCi-VSPの解明を行いたい。
【追加情報】(調査月2007年8月、記入月2008年6月)
関係論文
斎藤成也(国立遺伝学研究所)
増山和花(総合研究大学院大学生命科学研究科博士課程)
隅山健太(国立遺伝学研究所)
西野敦雄(日本学術振興会特別研究員)
岡村康司
電位センサータンパクが生物進化の過程でどのように変遷してきたかを明らかにするため,遺伝研側で,各 生物からのortholog遺伝子のアミノ酸配列をもとに,分子系統樹を作成した。解析の対象となった生物種は,ヒト,チンパンジー,カニクイザル,イ ヌ,ウシ,チキン,ゼブラフィッシュ,アフリカツメガエル (Xenopus tropicalis),ミドリフグ (Tetraodon nigroviridis),ホヤ,ウニ,オポッサム(Monodelphis domestica)と,アフリカツメガエル(Xenopus laevis),トラフグ (Fugu rubripes)であった。その結果,哺乳類の中では顕著に進化速度が速くなっていた。アミノ酸配列の比較により,電位センサードメインのS4の陽性 チャージの配列パターンが哺乳類とそれ以外で異なっていた。一方,岡崎側で,ヒト,マウス,ラット,トリ,ゼノパス,ゼブラフィッシュの各 ortholog分子を,ツメガエル卵母細胞に強制発現させ,電位センサー機能の電気生理学的計測を行った。その結果,ゼブラフィッシュ,ゼノパス,トリ では明確なゲート電流を示したが,哺乳類のVSPではゲート電流を出さなかった。この結果は分子系統樹での結果と良くあっており,哺乳類進化の過程で,膜 電位感知機能が変更され,生理的な役割も変化したことを示しているのかも知れない。今後は,1.哺乳類とトリの間に位置する生物種の分子機能解析も行 う,2.酵素活性についても種差を明らかにする,3.哺乳類のVSPにおいてアミノ酸配列を非哺乳類型に変更し,電位センサー機能が回復するかを調べ る,4.ウニやそれ以外の無脊椎動物のVSPを同定し,その分子機能を解析する,5.各動物種での組織発現パターンを比較する,などの解析を行い,生物進 化での変遷の意味を明らかにする予定である。
高橋 弘樹(基礎生物学研究所 形態形成研究部門)
山田 成宏(基礎生物学研究所 形態形成研究部門)
堀田 耕司(慶応義塾大学 理工学部生命情報学科)
小笠原 道生(千葉大学 理学部生物学科)
岡戸 晴生(東京都神経科学総合研究所 分子神経生理)
西野 敦雄(日本学術振興会特別研究員)
岡村 康司
神経発生過程における細胞間相互作用には生物種間で高度に保存された膜タンパク群の分子機能が重要であ る。しかし,これら膜タンパク質群の分子機能は遺伝子の点と点の繋がりとしては解明されているがダイナミックな時系列としての理解には程遠い。本研究で は,近年ゲノム情報が整備されるとともに単一細胞レベルでの個体発生が記述された原索動物ホヤ胚やEST情報が整備されたアフリカツメガエル胚を用い,神 経発生関連膜タンパク質とくにNotchシグナルや膜電位センサー分子のダイナミックな分子機能を個体まるごとのシステムで時系列的に解析を進めることを 目的とした。
1) 神経形成における脊索の役割
脊索動物の発生過程において,脊索は体軸伸長において働くのみならず,神経誘導や神経管形成過程におい ても重要な役割を果たしていると考えられているが,その分子メカニズムについては明らかにされていない点が多い。これまでに,我々はホヤのCi- Scale (Ci-Scabrous-like)のmRNAは脊索細胞のみに特異的に発現することを脊索動物で初めて明らかにしている。興味深いことにCi- Scaleタンパク質の局在をCi-Scale抗体を用いて調べたところ,脊索細胞および脊索細胞外の表皮感覚細胞および中枢神経細胞に沿った繊維状の構 造に局在することが明らかになった。
2) NotchシグナルによるScaleの局在変化
ショウジョウバエのScabrousは遺伝学的な解析からNotchと相互作用することが示唆されてい る。そこで,Ci-NotchとCi-Scaleの関係を解析するためにCi-Notch活性型の過剰発現あるいはガンマーセクレターゼ阻害剤による Notchシグナルを阻害したときのCi-Scaleの影響を調べた。その結果Ci-Notch活性化型を過剰発現させた場合は通常よりも多くのCi- Scaleが脊索外に局在するが,これとは逆に,Notchシグナルを阻害した場合は脊索外にほとんど局在しなかった。このことから,Notchシグナル によってCi-Scaleタンパク質の局在そのものが変化することが明らかになった。
3) Scaleの神経形成における機能解析
Notchシグナルを阻害すると同時にCi-Scaleの変異体を発現させると幼生期では,頭部におい て神経細胞の分化異常,尾部において神経軸索の走行異常が観察された。このことから,Ci-NotchとCi-Scaleは,神経細胞分化・形成において 協調的に働いていることが示唆されており,さらに機能解析を進めている。
宮脇 敦史(独立行政法人 理化学研究所 脳科学総合研究センター)
東島 眞一
cameleonは遺伝学的にコードされたレシオメトリックカルシウム指示薬である。最近,われわれ は,従来型よりもはるかに高感度にチューンされた次世代cameleonの開発に成功した。本研究では,イメージングに適した生物ゼブラフィッシュに次世 代cameleonをトランスジェニックゼブラフィッシュの手法により発現させて,様々な発生過程(たとえば体節形成)におけるカルシウムシグナルの関 与,および,(ii) 遊泳行動,逃避行動などの行動中にどのような神経細胞が活動するか,をカルシウムイメージングによって調べることを長期的な目標とした。まず,次世代 cameleon の一種,YC3.60を,すべての細胞で発現を促すhs70bプロモーターの下につないだコンストラクトを作製し,それに関してトランスジェニックフィッ シュを作製した。得られたトランスジェニックフィッシュでは,期待通りすべての細胞でYC3.60が発現しており,また,母性効果のためトランスジェニッ クフィッシュの母から生まれた胚では,1細胞期からYC3.60の発現が認められた。神経細胞で,その発火に伴うカルシウムシグナルが得られるかどうか を,感覚神経細胞であるRohan-Beard細胞を用いて検討した。ブンガロ毒素によって不動化した幼魚の皮膚に電気刺激を与えてシグナルの変化を見た が,期待に反して,明確なシグナルは認められなかった。これは,YC3.60は従来型のcameleonよりも高感度ではあるが,カルシウムとの親和性の 低いタイプのものであることが原因として考えられる。今後,カルシウムと高親和性タイプのYC2.60を用いて改めて検討を行う予定である。
小田 紀子(立命館大学 放射光生命科学研究センター)
山田 廣成(立命館大学 放射光生命科学研究センター)
山田 久夫(関西医科大学)
片岡 洋祐(関西医科大学)
富永 真琴(岡崎統合バイオサイエンスセンター)
恒温動物では体表温度は環境に依存して変動しているが,深部体温は常に37°C付近で保たれている。こ れは,体表の温度情報が神経を介して中枢の体温調節機構へフィードバックされているためである。近年,温度受容体タンパク質群(TRPV1~TRPV4) がクローニングされ,この4つのサブタイプTRPV1~TRPV4がそれぞれ異なる温度に反応して開閉するイオンチャンネルであること,またTRPV1は 辛味成分カプサイシンにも応答することが明らかとなった。しかし,体温付近の数度の温度変化を感知し,タンパク質の構造が変化する仕組みは未だ明らかでな い。そこで,TRPVの熱受容・体温調節機構への関与を検討するため,マウスに赤外線による熱刺激を与えたときの深部体温変化を測定した。
熱刺激に用いた赤外線は波長830nm,出力150mWの半導体レーザーで,皮膚温度が55度を超えな いように照射範囲を調節しながら5分間照射した。通常のマウスでは,この熱刺激で深部体温は変化せず,一定に保たれた。TRPV1に高濃度のカプサイシン を与えると脱感作が起こる。マウスに高濃度のカプサイシンを塗布し,TRPV1を脱感作させた後,同様の赤外線による熱刺激を与えると,通常では起こらな い体温上昇が計測された。この結果は,TRPV1の体温調節機構への関与を示すものである。
今後,TRPVの赤外応答の波長特異性について,近赤外から遠赤外に亘って強い放射光を発生する小型放 射光装置「みらくる20」からの放射光を分光照射して検討するとともに,数種類あるTRPVのうち特定のサブタイプの遺伝子欠損マウスを用いて,熱受容と 体温調節におけるTRPVサブタイプの役割を検討したい。
伊村 明浩(京都大学大学院医学研究科)
久保田 幸治(京都大学大学院医学研究科)
富永 真琴(岡崎統合バイオサイエンスセンター)
ヒト型老化疾患モデルマウスとして報告されたklotho(kl)マウスをヒントにし て,Klotho分子の機能を解析してきた。Klotho分子は一回膜貫通型のI型膜タンパクであり,主として遠位尿細管,脈絡膜,上皮小体の細胞内の膜 構造中に分布する。細胞膜表面に分布する量は少ないこと,また,膜貫通部位近傍で切断されて分泌され,体液中を循環することが判っていた。
脳脊髄液は中枢神経系を包み込んで灌流する体液であり,組成や圧力が厳密に制御されている。脈絡膜がその大部分を産生することは知られているが,その成分管理システムは知られていない。脳脊髄液中のCa濃度を測定したところ,kl-KO マウスでは有意に低下していた。そこで,脳脊髄液を感知するシステムに該当すると思われるセンサー分子を調べたところ,TRPV4が有力な候補の一つであ ると考えられるいくつかのデータを得た。Klotho分子はTRPV4シグナルに従って細胞膜イオン輸送体を細胞膜にリクルートするための分子装置である と推定して解析を進めている。
久保 伸夫(関西医科大学医学部)
沈 静(関西医科大学医学部)
小西 将矢(関西医科大学医学部)
濱田 聡子(関西医科大学医学部)
富永 真琴(岡崎統合バイオサイエンスセンター)
Ca2+ 透過性チャンネルと考えられている浸透圧感受性受容体(TRPV4)は,細胞内情報伝達機構において重要な役割を演じていることが明らかにされつつある。内耳液のイオン環境及び透過性の変化による局所の浸透圧変化は,蝸牛の外有毛細胞の運動能及び細胞内Ca2+ 濃度に影響を与え,結果聴覚機能になんらかの影響を与える可能性も考えられる。TRPV4の遺伝子が,蝸牛の内有毛細胞,外有毛 細胞,螺旋神経節細胞,血管条margin細胞に発現していることは既に報告されているがその機能は不明であった。そこで我々は,野生型及びTRPV4 knock-outマウスを用い,蝸牛のTRPV4の発現,単離外有毛細胞における低浸透圧刺激による細胞内Ca2+ 動態について比較検討を行った。
RT-PCR方法によって,野生型のマウス蝸牛においてTRPV4遺伝子の発現が検出された が,knock-out マウスの蝸牛では陰性であった。single-cell RT-PCR解析により,TRPV4遺伝子は野生型マウス蝸牛の螺旋神経節細胞,外有毛細胞,内有毛細胞に発現していることが確認された。これらの細胞で はTRPV4免疫反応も陽性であったが,knock-outマウスにおいては,いずれの細胞でも陰性であった。また,低浸透圧刺激及びTRPV4の agonistである4a-PDDに対して,野生型マウス外有毛細胞の[Ca2+]i上昇が認められた。この上昇はTRPV受容体のantagonistであるRuthenium Red により抑制された。一方,TRPV4 knock-outマウス外有毛細胞では,低浸透圧刺激及び4a-PDDによる[Ca2+]I 上昇が認められなかった。
以上の結果から,TRPV4受容体が外有毛細胞における細胞内情報伝達機構において重要な役割を演じている可能性が考えられた。
【追加情報】(調査月2007年8月、記入月2008年6月)
発表論文
野口 光一(兵庫医科大学医学部)
戴 毅(兵庫医科大学医学部)
富永 真琴(岡崎統合バイオサイエンスセンター)
TRPA1は2003年にクローニングされた新規チャンネルタンパクで,17度以下の侵害性冷刺激に よって活性化される。また,TRPA1はマスタードやマリファナの成分により活性化されることが報告され,痛みの標的分子の新しいメンバーとして注目を浴 びている。そこで,炎症疼痛モデル動物の疼痛過敏の発症メカニズムにおけるTRPA1の役割とその調節機構を明らかにすることを目的とする。TRPA1遺 伝子とタンパク質の発現を解析したところ,後根神経節細胞の約30% にTRPA1の発現を認めた。また,TRPA1は小径のTRPV1陽性細胞に発現していた。さらに,TRPA1が神経栄養因子受容体とも高率に共発現して いたことから,神経栄養因子のTRPA1活性への影響を感覚神経細胞においてCa2+イメージング法で検討したところ,神経栄養因子の投与は,リガンドによるTRPA1を介した細胞内Ca2+ 濃度上昇を増強した。よって,神経栄養因子はTRPA1機能を制御しているものと推測された。
【追加情報】(調査月2007年8月、記入月2008年6月)
発表論文
関係論文
宇理須 恒雄(分子科学研究所)
岡村 康司(岡崎統合バイオサイエンスセンター)
森垣 憲一(産業技術総合研究所)
内海 裕一(兵庫県立大学高度産業科学技術研究所)
富永 真琴(岡崎統合バイオサイエンスセンター)
イオンチャンネルの電気生理学的研究において,ピペット利用のパッチクランプが果たした貢献は計り知れ ないものがあるが,最近,新しいタイプのパッチクランプ:プレーナータイプのパッチクランプの研究が,特に欧米各国で活発化している。これは,1) 高集積が可能で,スクリーニング応用が可能,2) 二次元面内の多点測定が可能,3) 熟練を要しない,4) 小型により,体内挿入が可能,5) 蛍光や,細胞の活動電位の時空間分布との同時測定が可能,などの,ピペットパッチクランプに無い数々の特色があり,将来,マイクロ流体回路と組合わさった ものに取って代わることすら予測されてもいる。本共同研究では,シリコンを基板とするプレーナー型パッチクラン素子の開発を,富永が開発したTRPチャン ネル発現HEK293細胞を用いて進めている。素子開発における重要な技術的問題は,(1) 細胞あるいは,脂質ニ重膜を微細貫通孔に置いた時の孔のシール抵抗をギガオーム以上にすること(ギガオームシール),および(2)電流雑音を低減し,岡村 が研究を進めているプロトンチャンネルなどの微小電流チャンネルや高速応答計測を可能とすることである。本共同研究ではSi基板としてSOI構造基板を用 い,グラミシジンの系でシングルイオンチャンネルを計測し,雑音電流としてテフロン基板並みの低雑音を実現した。従来Si基板は雑音電流が大きいのでパッ チクランプの基板としては不適とされていた常識を変えるとことに成功した。
さらに微細貫通孔(径0.5-100mm)を 形成した基板をマイクロ流体回路に組み込み細胞を貫通孔部に導入し,現在,ギガオームシールを形成する実験をすすめている。今後,ギガオームシールの安定 な形成をめざした微細孔周辺の化学修飾,カプサイシンなどのリガンドを導入し,ホールセルモードや,シングルチャンネルの動作確認を行い,さらに,より低 雑音化,シール抵抗の安定高度化,マイクロ流体回路の高性能化などを進める。






