・私たち人間を含む真核生物の祖先に最も近縁な微生物を深海堆積物から培養することに世界で初めて成功した。
・培養した微生物は「アーキア」と呼ばれる微生物系統群に属し、他の微生物との共生に依存した生育をすること、真核生物に特有とされてきた遺伝子 (例えばアクチンやユビキチン等を作る遺伝子) を多く持つこと、細胞外に非常にユニークな触手状の長い突起を伸ばすこと等が明らかとなった。
・細胞構造や生理学的特徴ならびに生命史を踏まえ、真核生物の誕生について新しい進化説 (E3 model) を提案した。
国立研究開発法人海洋研究開発機構(理事長 松永 是)超先鋭研究開発部門の井町寛之主任研究員、国立研究開発法人産業技術総合研究所(理事長 中鉢 良治)生物プロセス研究部門のMasaru K. Nobu (延優)研究員らは、深海堆積物から真核生物の祖先に近縁な微生物(「アーキア(※1)に属する」)の培養に世界で初めて成功しました。
複雑な細胞構造を持つ真核生物 (人間、植物、魚類やカビ等) の祖先は、単純な細胞構造を持つ原核生物であるアーキアが、同じく原核生物であるバクテリアの細胞を内部に取り込み、共生させることによって誕生したとされています。共生したバクテリアはやがてミトコンドリアへの道を辿ります。最近の研究から、アスガルドと呼ばれるアーキアの一群が真核生物に最も近いと予測されていましたが、その存在は環境ゲノム情報でしか確認されておらず、その実体は完全に謎につつまれたままでした。
本研究では、独自のバイオリアクター技術 (2011年6月9日既報、2015年7月24日既報)、最新の遺伝子解析技術および従来型の培養技術を組み合わせて用いることにより、アスガルド群に属するアーキアの培養に挑戦しました。12年にわたる研究の結果、世界初のアスガルドアーキアの培養に成功しました。このアーキアにはギリシャ神話の神であるプロメテウスにちなみPrometheoarchaeum syntrophicum MK-D1株と名付けました (以下、MK-D1株という)。
MK-D1株はわずか直径550 nm (1 mmの約2000分の1) の球状細胞で、無酸素環境下でのみ生育しました。アミノ酸をエネルギー源として利用しますが、生育は他の微生物に依存することが明らかとなりました。ゲノム解析の結果、MK-D1株は真核生物に特有とされてきた遺伝子 (例えばアクチンやユビキチン等を作る遺伝子) を数多く保有していることが明らかになりました。このような遺伝子を保有しかつ真核生物に近縁であるにも関わらず、MK-D1株の細胞には真核生物のような複雑な細胞構造(例として核やゴルジ体)はなく、他のアーキアと同様に単純な内部構造であることが電子顕微鏡による詳細な観察結果から判明しました。その一方で、MK-D1株の細胞は他のアーキアやバクテリアでは観察されない複雑な形態を有することが明らかとなりました。特徴的であったのは、細胞外部に触手のような長い突起構造を形成することや多くの小胞を放出することでした。
さらに、本研究で培養に成功したMK-D1株の細胞構造や生理学的特徴およびこれまでの真核生物の起源に関連する研究結果に基づいて、「単純な細胞構造を持つ無酸素環境下で生育するアーキア」から「複雑な細胞構造を持ち酸素で呼吸する我々真核生物」への道筋を描いた真核生物の誕生についての新しい進化説”Entangle-Engulf-Endogenize (E3) model”を提案しました。
本成果は生物学の大きな謎である「真核生物の起源」について多くの洞察を与えるものです。今後、MK-D1株を使ったさらなる研究や、深海堆積物に生息するとされる真核生物に近縁な他のアーキアや原始的な単細胞の真核生物を培養することにより、アーキアから真核生物に進化した道筋がより詳細に明らかになっていくものと期待されます。
本研究の一部は、日本学術振興会の科研費JP18687006、JP21687006、JP24687011、JP15H02419、JP19H01005、JP18H03367、JP26710012、JP18H02426、JP18H05295、JP16J10845、JP18H04468、JP18K18795、先端バイオイメージング支援プラットフォームABiS (JP16H06280) および生理学研究所共同利用研究 (19-504) の支援を受けて実施されました。
本成果は、英科学誌「Nature」オンライン版に1月16日付け(日本時間)で掲載される予定です。
論文タイトル:
Isolation of an archaeon at the prokaryote-eukaryote interface
(原核生物と真核生物の境界に位置するアーキアの分離)
著者:井町寛之1*、Masaru K. Nobu (延優)2,1*、中原望1,2,3、諸野祐樹1、小河原美幸1、高木善弘1、高野淑識4、植松勝之5、生田哲朗6、伊藤元雄1、松井洋平4、宮崎征行1、村田和義7、齋藤由美1、酒井早苗1、Chihong Song (宋致宖)7、田角栄二1、山中結子1、山口隆司3、鎌形洋一2、玉木秀幸2、高井研1,8
1. 海洋研究開発機構 超先鋭研究開発部門、 2. 産業技術総合研究所 生物プロセス研究部門、 3. 長岡技術科学大学 環境社会基盤工学専攻、 4. 海洋研究開発機構 海洋機能利用部門、 5. 株式会社マリン・ワーク・ジャパン、 6. 海洋研究開発機構 地球環境部門、 7. 自然科学研究機構 生理学研究所、8. 自然科学研究機構 生命創成探究センター
*両著者はこの研究に同等に貢献しています。
我々、人間や植物を含む真核生物はどのようにして地球上に誕生したのでしょうか?真核生物の細胞はDNAを含む核、タンパク質と脂質膜からなる多数の小器官から構成される複雑な構造を持っています。この複雑な構造を持つ真核生物は、細胞内に核や小器官を持たない単純な細胞構造を持つ原核生物 (アーキアとバクテリア) から誕生したとされています。これまで真核生物細胞の誕生については様々な仮説が提案されてきましたが、最初の真核生物は原核生物であるアーキアが同じく原核生物であるバクテリア細胞 (ミトコンドリアの祖先) をその細胞内に取り込み、共生関係を築くことで誕生したという仮説が広く支持されています。しかしながら、これまで原核生物から真核生物に移行する中間体のような生物が見つかっていなかったため、その仮説の検証は困難であり、永らく真核生物の起源 についての研究は停滞していました。
しかしながら、2015年に状況は大きく変わってきます。スウェーデンの研究チームが、メタゲノム解析(※2)を用いて深海堆積物に生息するロキアーキオータと呼ばれる未培養アーキア群のゲノムを解読し、分子系統解析からこのアーキア群が真核生物に最も近縁であると主張しました。その主張を支持するように、ロキアーキオータのゲノムにはこれまで真核生物に特有とされてきた遺伝子が多数存在することがわかりました (例えば真核生物細胞の骨格形成に関わるアクチンや小胞体輸送に関連する遺伝子等)。さらに同研究チームはロキアーキオータに近縁な別のアーキア群も真核生物に特有とされてきた遺伝子を多数保有していることをメタゲノム解析により明らかにしました。ロキアーキオータとその近縁なアーキア群をまとめた分類群 (上門) は、北欧神話の神々が住まう世界にちなんでアスガルド上門と名付けられ、現在、このアーキア群が真核生物起源の有力候補と考えられています。しかしながら、本アーキア群は環境ゲノム情報でしか存在が確認されていないため、その実体 (細胞形態や生き様等) は完全に謎に包まれたままでした。本アーキア群が本当に真核生物の起源たる生物であるのであれば、例えば、①真核生物と同様に核や小器官を有する複雑な細胞構造をすでに有しているのか、②ミトコンドリアの祖先となるバクテリアを細胞内に取り込むことができたのであれば、細胞外部から内部へ大きな物質を取り込む能力である食作用 (ファゴサイトーシス) を有しているのか、という問いが自然と湧いてきます。これらの問いに答えるためには、アスガルド上門に属するアーキアを実験室で培養して詳細に調べることが必須となります。そのため、アスガルド上門アーキアの培養が世界的に大きく期待されていました。
一方で、本論文の筆頭著者である井町は、アスガルド上門アーキアが真核生物の起源として注目され始める2015年よりもずっと以前の2006年からその培養に挑戦していました。その理由は、アスガルド上門の1つであるロキアーキオータは世界中の海底堆積物に優占的に存在していることが16S リボソームRNA遺伝子(※3)に基づいた遺伝子解析から知られており (当時はロキアーキオータという名前ではなくDeep-Sea Archaeal Group [DSAG] あるいはMarine Benthic Group-B [MBG-B] と呼ばれていた)、このアーキアを培養することで、その生理学的特徴の解明や環境中での役割を解明したいと考えていたからです。
本研究では、2006年5月に有人潜水調査船「しんかい6500」により、ロキアーキオータを含めた多種多様な未培養微生物を含む深海堆積物を、南海トラフのメタン湧出帯 (水深2533 m) から採取しました (図1)。採取した深海堆積物は下降流懸垂型スポンジ (down-flow hanging sponge: DHS) リアクターに入れ、メタン湧出帯の環境を模擬した培養条件により約2000日間の培養を行いました。ロキアーキオータを含む多種多様な未培養微生物がDHSリアクター内に培養されていることを16S リボソームRNA遺伝子に基づいた遺伝子解析を用いて確認しました。続いて、様々な培地を入れた試験管を数百本準備し、DHSリアクター内に培養されてきた個々の微生物の分離を試みました。約1年後に、カザミノ酸 (※4)と抗生物質 (バクテリアの生育を抑えるため) を加えた嫌気培地が入った試験管において、微生物の増殖と思われた微弱な培地の濁りを観察しました。その培養物を16S リボソームRNA遺伝子に基づいた遺伝子解析を用いて調べた結果、少数のロキアーキオータとその他数種類の微生物が存在することが判明しました。その後、本ロキアーキオータの増殖を定量PCRでモニタリングしながら、培養物の植え継ぎを行いました。その過程で、本ロキアーキオータの増殖速度は非常に遅いこと (細胞の倍加時間(※5)は約14〜25日)、そして増殖時の最大細胞密度が極度に低いこと (〜105 cells/ml) が明らかとなりました。この増殖速度と最大細胞密度は、最もよく研究されているモデル微生物の大腸菌と比較して1000倍程度低いものでした。さらに、FISH法、電子顕微鏡観察、メタゲノム解析および安定同位体標識アミノ酸を用いた培養実験により、本ロキアーキオータは無酸素環境下でアミノ酸を代謝し、その代謝産物として生成される水素を他の微生物 (硫酸還元バクテリアやメタン生成アーキア) に渡すことで共生関係を維持しながら増殖していることが明らかとなりました。最終的に、深海堆積物を採取してから12年後に、本ロキアーキオータをMethanogenium属メタン生成アーキアとの2種類の微生物からなる純粋な共培養系として分離することに成功しました。本ロキアーキオータはギリシャ神話の神であるプロメテウスが泥から人間を作り、人類に火を与えたとされる(ミトコンドリアを獲得し酸素が利用できるになったことで莫大なエネルギーを利用できることになった)ことからPrometheoarchaeum syntrophicum MK-D1株と名付けました。
MK-D1株は無酸素環境でのみ増殖可能な嫌気性微生物であり、アミノ酸あるいはペプチドをエネルギー源として利用し生育できることがわかりました。各種電子顕微鏡を用いて詳細な細胞構造を調べた結果、MK-D1株の細胞は約550 nm (1 mmの約1/2000) の直径を持つ小さな球菌であることや増殖時には細胞同士が集まった凝集体を形成して生育することが明らかとなりました。さらに細胞の内部構造をクライオ電子顕微鏡トモグラフィーや透過型電子顕微鏡で調べた結果、他のアーキアと同様に、MK-D1株の細胞内部は単純な構造で、多くの研究者が予測していた「アスガルド上門アーキアは真核生物と同様に核や小器官を有する複雑な細胞である」ことを裏切るものでした。その一方で、MK-D1株の細胞形態は他の原核生物では観察されない複雑な形態を有することが明らかとなりました。特筆すべき特徴は、触手のような長くて分岐を有する突起構造を細胞外部に形成することでした(図2)。さらに細胞外には多くの小胞を放出することもわかりました。細胞の進化を考える上で重要な細胞膜を構成する膜脂質分子の調査も行いました。その結果、MK-D1株は、これまでに知られているアーキアと同様にイソプレノイド型の膜脂質分子構造を有していました。現存する真核生物の細胞膜は脂肪酸がエステル結合した構造であることを考えると、アーキアが真核生物細胞に進化していく過程で、細胞膜脂質がイソプレノイド型から脂肪酸型への切り替えが起きたと推定されます。
今回MK-D1株を培養することができたため、MK-D1株の完全長ゲノム配列を決定することができました。本完全長ゲノム情報を用いて、リボソームタンパク質(※6)配列に基づく分子系統解析を行った結果、MK-D1株は真核生物群と姉妹群を形成したことから、「MK-D1株は培養された原核生物として真核生物に最も近縁な生物である」ことが示されました (図3)。加えて、MK-D1株のゲノムにも真核生物に特有とされてきたアクチン、ユビキチンや小胞体輸送関連の遺伝子が多数存在しており、RNA発現解析からそれらの遺伝子群が実際に細胞内で発現していることも確認しました。それら真核生物に特有とされてきた遺伝子の中ではアクチンを作る遺伝子が最も多く発現していました。さらにMK-D1株のゲノムを詳細に解析したところ、自身の細胞を作るために必須なアミノ酸、ビタミン類やヌクレオチドを自身では合成できないことが示唆されました。この結果は、MK-D1株がエネルギー源であるアミノ酸の利用を他の微生物との共生に依存しているだけでなく、自分自身の細胞合成や生育に必須となる物質も他の微生物からの供給に依存していることを示唆しています。さらに、MK-D1株のゲノムとこれまでに報告されているアスガルド上門アーキアとのゲノム情報の比較を行ったところ、アスガルド上門アーキアにおいては、アミノ酸からのエネルギー獲得に関連する遺伝子が保存されていることがわかりました。この比較ゲノム解析の結果から、アスガルド上門の祖先となるアーキアはアミノ酸を利用して生育していたことが強く示唆されました。従って、真核生物の祖先となるアーキアはアミノ酸をエネルギー源として利用して生育していたと考えることができます。
最後に、本研究で明らかになったMK-D1株の細胞形態・生理・遺伝学的な特徴とこれまでの真核生物の起源に関連する研究結果に基づいて、我々は真核生物の誕生についての新しい進化説”Entangle-Engulf-Endogenize (E3) model”を提案しました (図4)。今から約27億年前、シアノバクテリアの登場により地球に酸素が増えてくる大酸化イベントが始まりました。真核生物の祖先となるアーキアは、彼らにとって毒である酸素を解毒してもらうためにミトコンドリアの祖先となるバクテリア細胞と共生します。そして、そのアーキアはMK-D1株で観察されたような長い触手や小胞を使ってミトコンドリアの祖先を取り込みます。最終的に、アーキアとミトコンドリアの祖先との共生関係が進行し成熟した結果として、最初の真核生物細胞が生まれたという仮説です。
まずはMK-D1株についてさらに詳細な調査を進めていく予定です。アーキアであるMK-D1株が持つ真核生物に特有とされてきた遺伝子の働きを調査することで、それら遺伝子の本来の役割を理解することができる可能性があります。また、MK-D1株で観察された触手状の長い突起構造や小胞の構成成分やその役割の解明を目指します。さらに、MK-D1株以外の真核生物に近縁とされている他のアーキアや、原始的な単細胞の真核生物の培養を試み、その性質も明らかにしていく予定です。これらの研究を進めることによりアーキアから真核生物に進化した道筋がより明らかになっていくものと期待されます。
※1 アーキア:アーキアは生物界の主要な系統群の1つであり、バクテリア (真正細菌) と真核生物と共に全生物界を3分している存在である。古細菌とも呼ばれる。アーキアはすべて人間の目で直接見ることができない微生物であり、細胞内に核を持たない原核生物である。バクテリアも同様に原核生物であるが、細胞膜の組成や構造等に違いがあり、全く別の生物である。アーキアは高温やpHの低い極限的な環境にしかいないと以前は考えられてきたが、現在では我々の身の回りの環境にも普遍的に存在していることが遺伝子解析技術の発達により明らかにされている。
※2 メタゲノム解析:環境試料から直接回収したゲノムDNAを塩基配列決定装置により網羅的に解読する手法のこと。環境中の多くの微生物が培養困難であるため、現在の微生物学では、微生物の代謝機能を予測する手法として広く用いられている。
※3 16S リボソームRNA遺伝子:遺伝子の翻訳の場であるリボソームを構成するRNAの1種をコードしている遺伝子。リボソームおよびそこに含まれるRNAはすべての生物に存在しており、16S リボゾームRNA遺伝子は進化速度が比較的遅いため、その遺伝子配列情報は生物の系統分類に広く用いられている。また、環境試料に含まれる微生物の種類や培養されてきた微生物の種類を調べるために、抽出したDNAを鋳型として16S リボゾームRNA遺伝子部位をPCRにより増幅して調べる手法はメタゲノム解析が普及する以前から広く利用されている。
※4 カザミノ酸:カゼインを酸で加水分解したものであり、アミノ酸やペプチドの混合物である。ただし、酸で加水分解を行うためにトリプトファンは破壊されてしまい、含まれていない。微生物の培地成分の1つとしてよく使われる。
※5 倍加時間:1つの細胞が2つの細胞に分裂するまでの時間。最も研究されている微生物の代表である大腸菌の倍加時間は約20分。
※6 リボソームタンパク質:遺伝子の翻訳の場となっているリボソームを構成するタンパク質の総称。リボソームタンパク質配列を連結させて分子系統解析を行うと、より精緻な生物間の関係が理解できるとされている。
(本研究について)
国立研究開発法人海洋研究開発機構
超先鋭研究開発部門 超先鋭研究プログラム
主任研究員 井町 寛之
国立研究開発法人産業技術総合研究所
生物プロセス研究部門 生物資源情報基盤研究グループ
研究員 Masaru K. Nobu (延 優)
(報道担当)
国立研究開発法人海洋研究開発機構
海洋科学技術戦略部 広報課
国立研究開発法人産業技術総合研究所
企画本部 報道室
国立大学法人長岡技術科学大学総務部大学戦略企画・広報室
大学共同利用機関法人自然科学研究機構生理学研究所
研究力強化戦略室
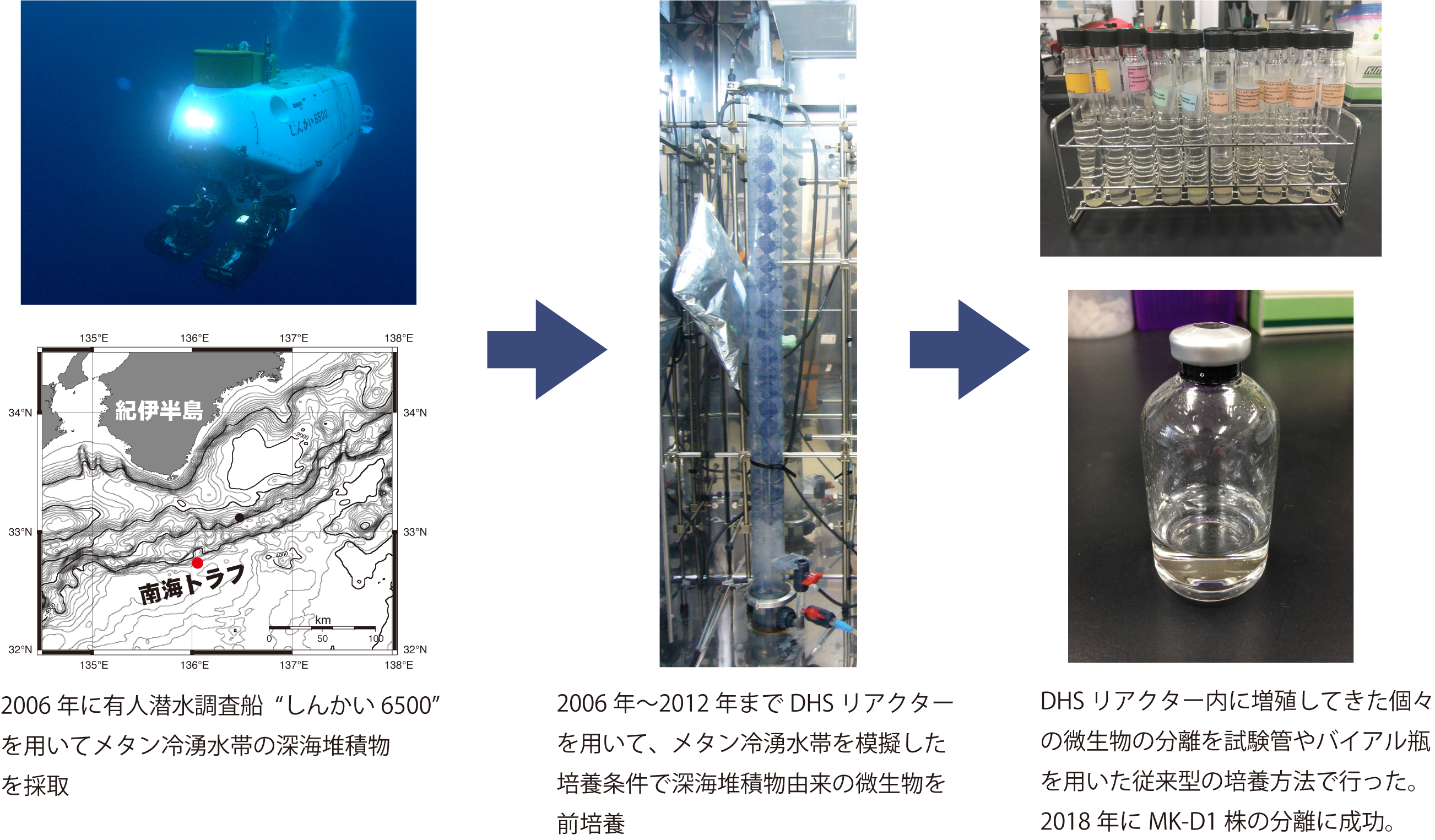
図1 深海堆積物からMK-D1株を分離するまでの過程。
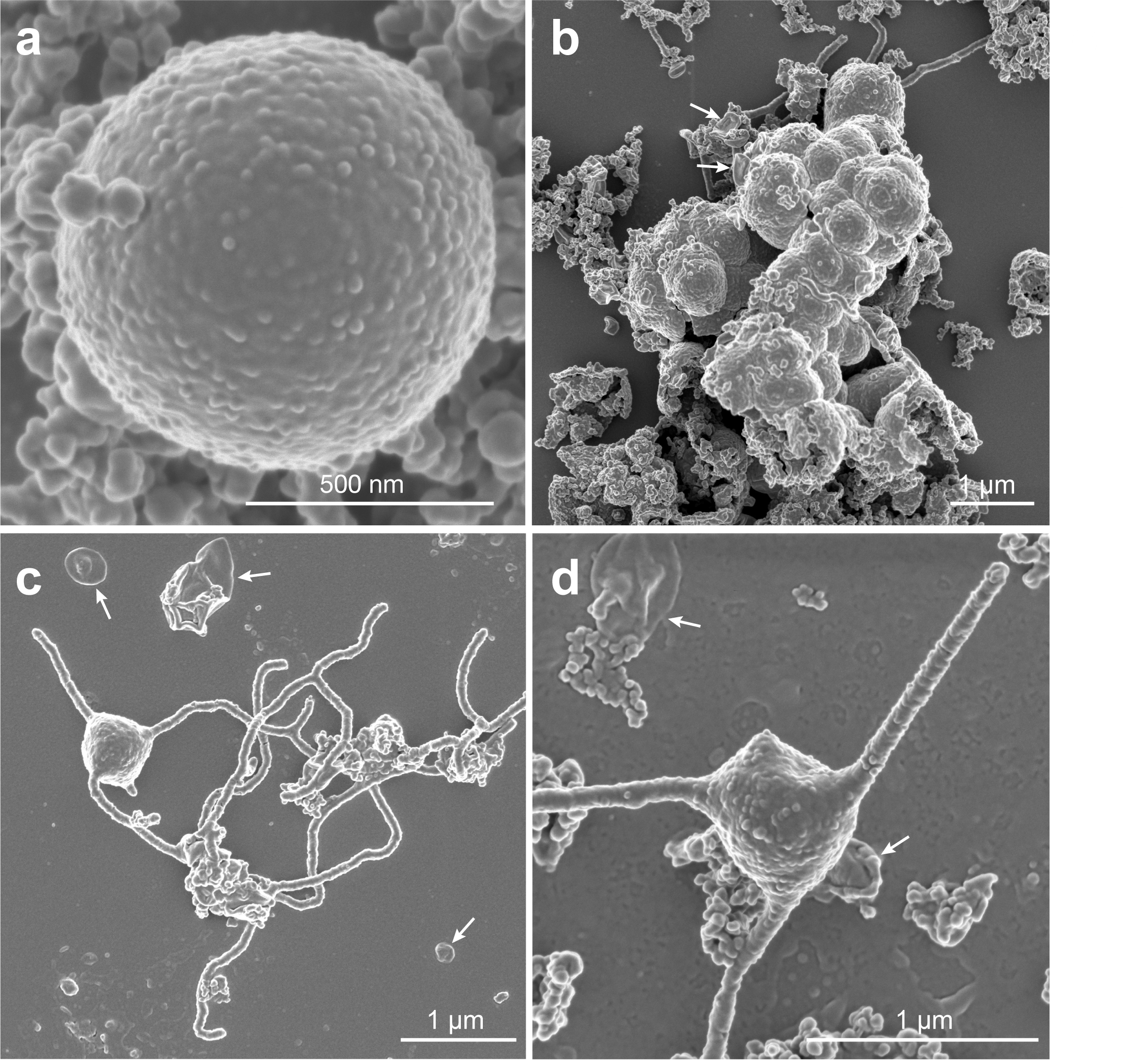
図2 培養したアーキアMK-D1株の走査型電子顕微鏡写真。上2つの写真は、増殖しているときの細胞形態。単独、あるいは多くの場合で集塊体を作っている。下2つの写真は、増殖が終わる頃の細胞形態で、触手状の長い分岐した突起を作る。白い矢印は、MK-D1株が細胞外に放出した小胞。
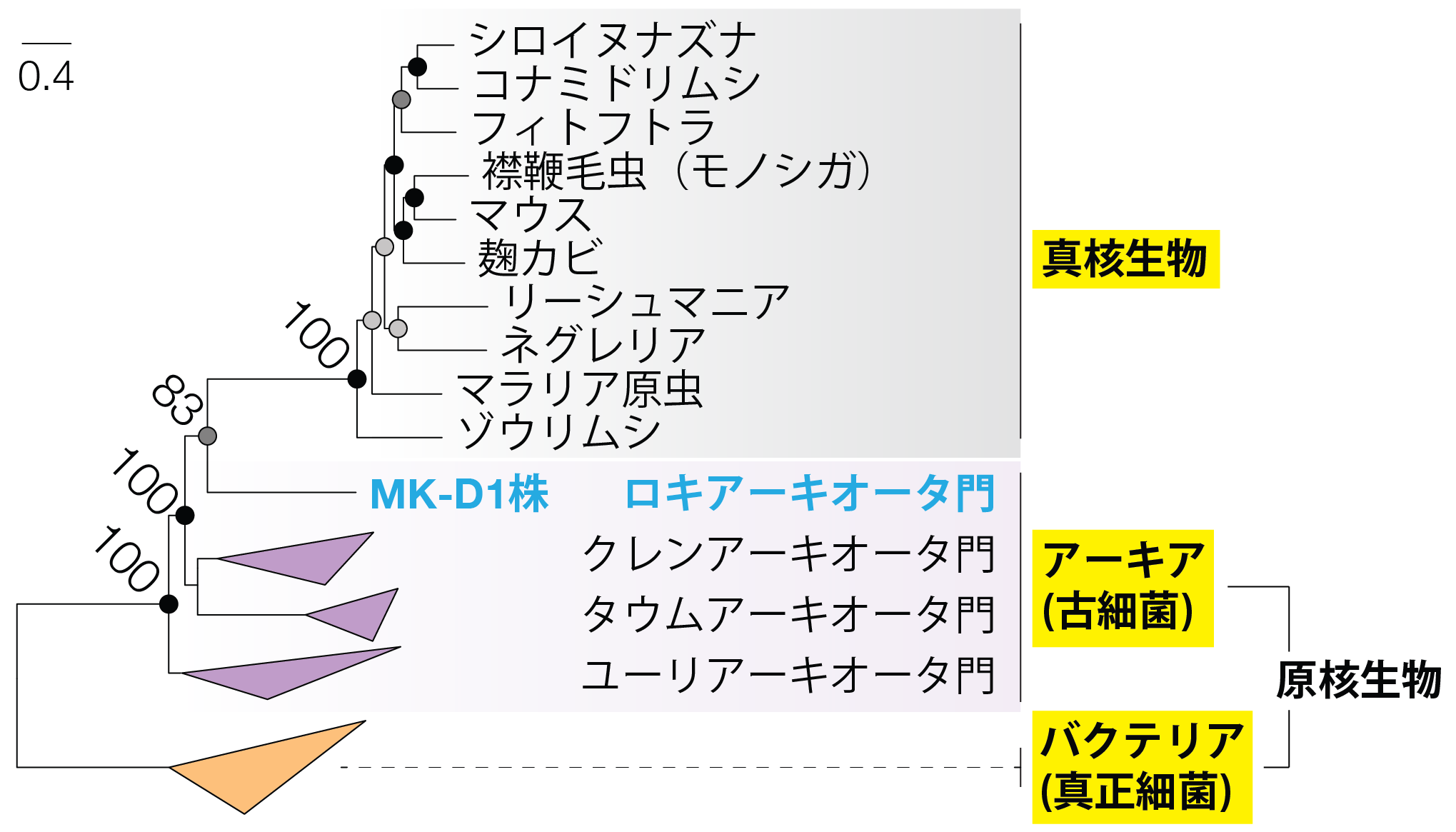
図3 リボソームタンパク質配列を連結した配列を用いて最尤法により構築した分子系統樹。MK-D1株は培養された原核生物として真核生物に最も近縁な生物であることがわかる。枝の分岐点にある数字はそこの分岐の確率(%)を示す。横方向の長さは進化的な距離を示している(スケールバーは左上を参照)。
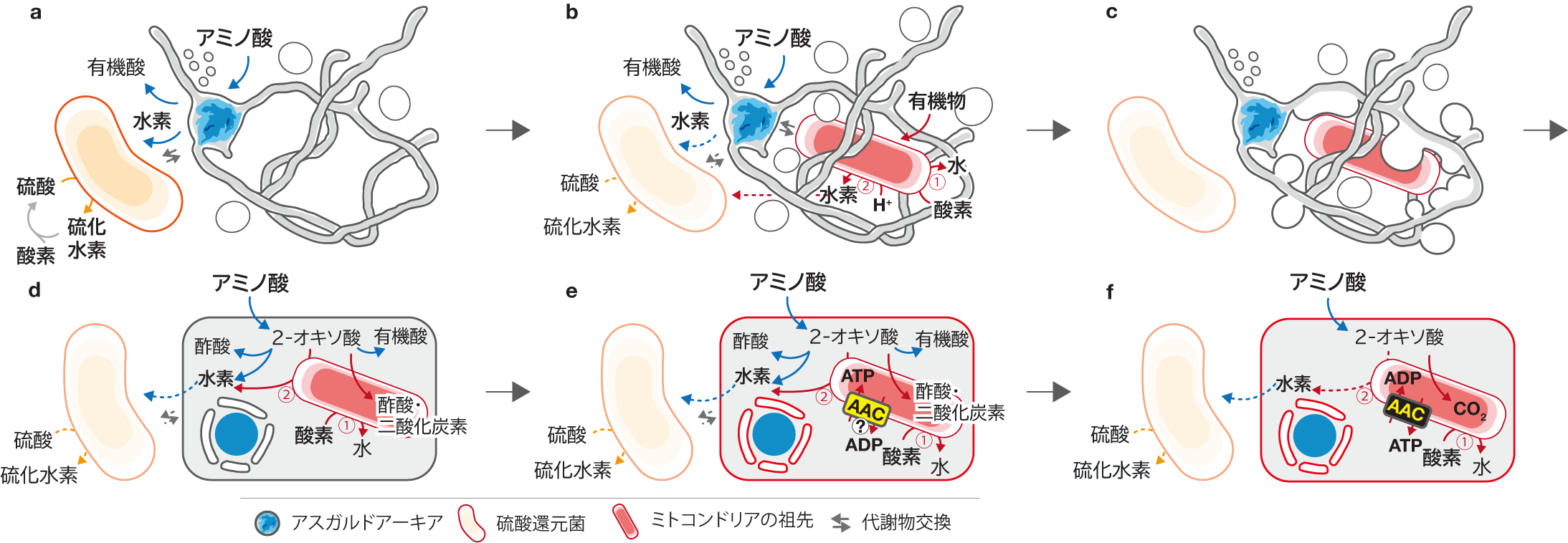
図4 提案した真核生物の誕生についての新しい進化説”Entangle-Engulf-Endogenize (E3) model”。Entangleは巻き込む、Engulfは飲み込む、そしてEndogenizeは内部に発達させるという意味であり、真核生物の祖先となるアーキア(青色で示した細胞)がミトコンドリアの祖先となるバクテリア(赤色で示した細胞)を細胞内に取り込み、その共生関係が成熟していくそれぞれの過程を表現していている。約27億年前に始まった大酸化イベントによって酸素が生成されるようになると、無酸素環境でのみ生育可能な真核生物の祖先となるアーキアは酸素存在下でも生育できる硫酸還元菌(オレンジ色で示した細胞)や酸素を消費・解毒できるバクテリア(ミトコンドリアの祖先)と共生するようになった。そしてアーキアが持つ触手状の突起と小胞により酸素を利用できるバクテリア細胞はアーキア細胞に取り込まれた。1つの細胞になったアーキアとバクテリアはアミノ酸を分け合うことで共存した。アーキア自身の「殻(細胞壁)」があったお陰で膜が完全に閉じられず、アーキア本体とミトコンドリアを分ける「核膜」が形成された。共生関係が発達していく過程で遺伝子交換が行われ、さらに一体化が進んでいった(細胞を囲む膜がアーキア型からミトコンドリアと同質になる等)。ミトコンドリアの祖先が宿主アーキアのエネルギー通貨(ATP)を奪う機能(AAC)を作り発達させることで、アーキアとバクテリアの関係が共生から寄生に切り替わった。アーキアはエネルギー生産代謝をミトコンドリアに完全に委ねることによりAACの逆流を促し、ATPを二者で共有する共生システムを確立した。この一連の進化を通じて、アミノ酸をエネルギー源、酸素で呼吸するシステム、核膜等の複雑な細胞構造を有する私たちの祖先である真核生物が生まれた。
![]()
![]()
![]()
![]()
![]()
国立研究開発法人海洋研究開発機構
国立研究開発法人産業技術総合研究所
国立大学法人長岡技術科学大学
株式会社マリン・ワーク・ジャパン
大学共同利用機関法人自然科学研究機構






