斉藤修,伊藤政之(長浜バイオサイエンス大学,バイオサイエンス学部)
久保義弘
我々は,G蛋白質共役応答の調節因子としてGaGAPの作用をもつRGSタンパクファミリーに注目して,RGS8とRGS8S(N端部9残基のみが異なる)をクローニングして解析を行ってきた。そして,Gq受容体シグナルの制御能を解析した結果,RGS8が,Gaq とは結合性が低いにもかかわらず,各ムスカリンGq受容体に対して受容体選択的なGq抑制能を示し,M1系は抑制するが,M3系の抑制は弱いこと,さらに RGS8SはいずれのGq系にも制御能が弱いことなどが判明した。そこで,このRGS8の受容体選択的なGq調節の機構として,RGS8が直接特定の受容 体に選択的に結合している可能性を検討した。
まず,受容体との直接の反応性を組換え蛋白質を用いた共枕実験で解析した。すると,RGS8がGq共役 のM1とM3のムスカリン受容体の第三細胞内ループ (i3) に結合することが判明し,特にM1へは高い結合性が検出された。一方,Gi共役のM2受容体のi3へは,RGS8は全く結合しないことが明らかになった。 また,RGS8Sについては,いずれの受容体への結合性もRGS8に比べ弱く,それらの結合性は両RGS8分子の示すGq制御の選択性とよく一致すること が判明した。そこで,次にそのRGS8と受容体との直接の相互作用が実際の細胞内で起こっているのか,BRET (Bioluminescence Resonance Energy Transfer) 解析を行った。すると実際の細胞では,RGS8はM1に結合しているのに対して,RGS8Sは殆ど結合していないことが明らかになった。次に,M1のi3 のどの部位にRGS8が結合性を示すのか,i3を3パートに分け検討した。結果,RGS8と8Sが共通に認識する領域が細胞内ループのN端側に,RGS8 が特異的に結合する部位がC端側50残基に存在することが示された。また,RGS8のN端側からの欠失変異体を作成して,RGS8が受容体を認識する部位 をサーチした。結果,RGS8のN端6~9残基のわずか4残基の配列「MPRR」がM1受容体認識に必須な部位であることが判明した。そこで,この配列の 特に連続したR残基に注目してその両残基をAに置換した変異体を作成したところ,M1i3への結合能が激減することが明らかになった。以上のように RGS8の受容体選択的なGq調節の機構と考えられる特定の受容体への直接結合とその分子的基盤が明らかになってきた。
今後は生理学的解析によって,このRGS8の受容体への直接結合が如何に受容体選択的なGq調節能に関わっているのか明らかにしていきたい。
【追加情報】(調査月2007年8月、記入月2008年6月)
発表論文
柳(石原)圭子,鄢定紅(佐賀大学医学部)
上原明,中村友紀(福岡大学医学部)
Batu KECELI(Hacettepe 大学医学部,トルコ)
久保義弘
イオンチャネルやイオン透過型受容体は膜電位変化やリガンド結合が引き金となり分子構造が変化すること によって細胞膜のイオン透過性を変化させ,様々な細胞機能の発現に関与する。内向き整流カリウムチャネルは神経や心筋細胞等の膜興奮性の調節をつかさどる イオンチャネルであり,7つのサブファミリーに属する多くの種類がある。私たちは内向き整流カリウムチャネルの分子構造と機能の連関を探る目的で,今年度 は強い内向き整流性カリウムチャネルのイオン透過孔を形成するサブユニットであるKir2.1 (IRK1)とG蛋白制御内向き整流カリウムチャネルのイオン透過孔を形成するサブユニットであるKir3.4(GIRK4)がヘテロ多量体を形成するか どうかについて着目し実験を行った。
まずHEK293細胞に共発現させたIRK1とGIRK4の結合の有無をFLAGあるいはmycタグを 結合したIRK1あるいはGIRK4を用いて免疫共沈法で調べたところ,IRK1-FLAGとGIRK4-mycおよびGIRK4-FLAGとIRK1- mycのペアは共に陽性コントロールであるIRK1-FLAGとIRK1-mycのペアと同程度に共沈するという結果を得た。そこでアフリカツメガエル卵 母細胞を発現系として用いて電気生理学的検討を行ったところ,IRK1とGIRK4のヘテロ多量体がチャネルとして機能するという証拠は得られなかった が,IRK1のdominant- negative型変異体(ポア領域GYGをAAAに変換)がGIRK4とGIRK1から成るチャネルを流れる電流を有意に減少させるという結果を得たた め,IRK1とGIRK4(とGIRK1)がヘテロ多量体を形成することが示唆された。さらにCFPあるいはYFPを結合したIRK1あるいはGIRK4 を作製し,FRET(蛍光共鳴エネルギー転移)解析を行ったところ,これらがヘテロ多量体を形成していることを示唆する結果を得た。以上の結果は異所性発 現系においてIRK1とGIRK4がヘテロ多量体を形成し得ることを示した (Journal of Physiological Sciences 56: S151, 2006)。今後はIRK1とGIRK4が共発現している心房筋細胞においてヘテロ多量体が存在しているかどうかを含め,さらにその詳細を明らかにした い。
【追加情報】(調査月2007年8月、記入月2008年6月)
発表論文
関係論文
和田洋巳,田中文啓,片倉浩理,佐久間圭一郎長谷純宏,長束俊治(大阪大学大学院理学研究科),中北愼一(香川大学総合生命科学実験センター)
東海林博樹(香川大学)山中龍也,土屋尚人(新潟大学脳研究所)
辻崇一(お茶の水大学大学院糖鎖科学研究センター)鈴木邦彦(東海大学未来科学技術共同研究センター)
N-結合型糖鎖は主に細胞表面に存在し,他の細胞や細胞外基質との相互作用を介して様々な機能を果たすことが知られている。我々はこれまでにマウスの正常発達過程やヒトの癌,神経変性疾患などを対象にN-結合型糖鎖発現パターン解析を行い,糖鎖の多様な機能の解明を目指してきた。
シアリルルイスX構造をはじめとして,糖鎖が癌転移に大きく関与することは既に明らかであるが,糖鎖に 関して遺伝子やタンパク質のような網羅的発現パターン解析を行った研究は非常に少ないため,糖鎖機能の多くが未解明であることが予想される。我々は独自に 樹立した定量的糖鎖パターン解析系を用いて,癌の肺転移において特に重要な糖鎖構造の特定を行った。
マウスにおいてB16(マウスメラノーマ細胞)を用いて肺転移と肝転移モデルを作成し,両者の糖鎖パ ターンの比較からA2G2F構造が肺転移に特に多いことを見出した。また,B16の中でA2G2F構造を多く持つ細胞集団は少ない集団に比べてより肺転移 しやすいことを見出した。さらに,臨床剖検標本を用いて肺転移と肝転移のLCA (Lens culinaris agglutinin) 染色を行い,ヒトにおいてもA2G2F構造が肺転移を促進している可能性を示した。(Biochem Biophys Res Commun. 2006;340(3):829-35)
特定の糖鎖構造が臓器選択的転移に関与することを示した既報は無く,この研究によって癌転移機序の新たな一面が示される可能性があり,非常に興味深い結果といえる。
【追加情報】(調査月2007年8月、記入月2008年6月)
発表論文
関係論文
馬場広子,山口宜秀,林明子,鈴木彩佳,星登美子,高橋ゆかり(東京薬科大学薬学部)
渡辺修一,中平健祐(埼玉医科大学)中平英子,長谷川明子(国立精神神経センター神経研究所)
村上志津子(順天堂大学医学部)清水健史(熊本大学発生医学研究センター)浜崎浩子(東京医科歯科大学)
神経細胞の発生や機能分化に対しては,神経細胞自体の内在性要因の他に,周囲の細胞や細胞外基質からのシグナルが重要な役割を果たしている。本研究では,神経細胞の興奮性の獲得に関わる細胞内外のシグナルの分子機構を明らかにすることを目的とし,下記の結果を得た。
1) パラノーダルジャンクション形成不全に伴う遺伝子発現変化の解析
スルファチド欠損マウスでは,軸索・ミエリン間のパラノーダルジャンクション (PJ) の形成不全を生じる。この結果,正常ではランビエ絞輪周辺に集積する電位依存性イオンチャネルの局在が加齢と共に変化し,それに伴ってふるえなどの神経症 状を呈する。このマウスの脊髄では脱髄性変化はほとんど生じないにもかかわらず,軸索表面のチャネル変化と同じ時間的経過で著明なアストロサイトの活性化 が生じる。そこで,明かな軸索変化を生じている週齢のマウス脊髄mRNAを抽出し,マイクロアレイ法を用いて発現が変化する遺伝子の解析を行った。結果, スルファチド欠損で発現が上昇する遺伝子および低下する遺伝子が複数得られた。これらの遺伝子の発現細胞およびその機能と軸索変化に対する関連について現 在検討を行っている。
2) 電位依存性Na+チャネル発現ベクターの構築有髄神経軸索では,ミエリン形成に伴って電位依存性Na+チャ ネルのサブタイプの変換と局在部位の変化を生じる。無髄の状態ではNav1.2が軸索に存在するが,ミエリン形成によってランビエ絞輪ができる頃に Nav1.2からNav1.6へとサブタイプの変換がおこる。しかし,その変換の機構に関してはまだ不明である。PJ形成不全マウスではこの変換が不完全 であることから,PJ形成がサブタイプ変換に対して何らかの影響を与えていると考えられる。そこで,Nav1.6の軸索への輸送や局在化の機構を明らかに する目的で,マウスNav1.6のcDNA全長をRT-PCRでクローニングした。現在発現ベクターを構築中である。
3) スルファチド欠損に伴う末梢神経系ランビエ絞輪の変化の解析
スルファチド欠損マウス末梢神経系におけるランビエ絞輪部の変化を調べた。免疫組織染色では大きな変化 を認めなかったが,電子顕微鏡を用いた解析では,微絨毛および絞輪軸索部分の明らかな形態変化を認めた。スルファチド欠損あるいはそれに伴うミエリン異常 とこの絞輪軸索の異常との関係を現在解析中である。
【追加情報】(調査月2007年8月、記入月2008年6月)
発表論文
関係論文
清水惠司,中林博道,朴啓彰,豊永晋一,八幡俊男,土屋孝弘,政平訓貴,中居永一(高知大学医学部)
栗山茂樹,峠哲男,出口章広(香川大学医学部),中根恭司(関西医科大学),中村秀次(兵庫医科大学)
悪性グリオーマは,放射線療法,化学療法および免疫療法の著しい開発をもってしても,ここ数十年来,治 療成績の大きな向上が得られていない難治性悪性腫瘍である。その原因には,この腫瘍の高い浸潤能,各種療法に対する耐性能の獲得が挙げられる。それ故,こ の腫瘍を根治するためには,既存の治療法とは質の異なる革新的な新規治療法の開発が必要とされるのは明白である。
我々は,現在までに,この疾患に対するレトロウイルスベクターをもちいた自殺遺伝子療法を開発してきた。パッケージング細胞の遺伝的改変と遠心濃縮により1x1011-12 pfu/ml の高力価ウイルス溶液の調製を可能とした。さらに,自殺遺伝子 HSVtkの発現制御にミエリン塩基性蛋白遺伝子のプロモーターを導入し,脳特異的高力価レトロウイルスベクターを構築した。これを用いた遺伝子治療にて,ガンシクロビル投与プロトコールの最適化をおこない,マウスグリオーマモデルを完治せしめた。
脳特異的高力価レトロウイルスベクターを臨床応用するために霊長類(コモン・マーモセット)を用いた毒性・発ガン性試験を開始した。患者に一回あたり投与する予定量の1x1011 pfu のレトロウイルスをコモン・マーモセットの脳実質に投与し,経過観察をおこなった。3ヶ月後の血液検査,8ヶ月後のMRIによる撮像,血液検査及び解剖に おいて異常所見は観察されなかった。各組織からのウイルスゲノムの検出をPCR法にてDNA,RNAレベル共に実施した結果,残存ウイルスは検出されな かった。さらに,レトロウイルスベクターをもちいる際に問題となるRCR (Replication Competent Retroviruses) の有無の試験をFDA(米国食料医薬品局)の検査基準でおこなえる外部機関に委託した結果,RCRの混入は陰性である証明を得た。今後は,安全性試験の検 体数を増やし,臨床試験の準備に着手する予定である。
現在,我々は脳特異的高力価レトロウイルスベクターの前臨床段階試験に加え,安全性を考慮した治療用ベ クターを構築中である。これらの開発により,悪性脳腫瘍に限局せず,他組織由来の悪性腫瘍,他の疾患に有用な安全で効果の期待できるウイルスベクターを提 供すると共に悪性脳腫瘍に対する遺伝子治療法の確立を目指している。
【追加情報】(調査月2007年8月、記入月2008年6月)
発表論文
関係論文
河西春郎(生理研)
阿部輝雄(新潟大学脳研究所)
白尾智明(群馬大学医学部)
大脳皮質の錐体細胞樹状突起のスパインは2光子励起観察法やGFP導入の導入により,形態や更にその機 能の変化の長期的追跡が可能となってきている。我々は,更に,2光子励起グルタミン酸法によって,スライス培養標本において単一スパインの形態可塑性を誘 発することを可能とし,これに機能変化が伴うことを見出している。シナプス可塑性に関わる分子・超分子のダイナミズムが可視化定量化するための分子標識 法,有効な追跡分子,標本の選択などについて検討を行ってきた。本年度は,2光子活性化が可能なGFP (PAGFP) をアクチンに融合させたPAGFP-actinを用いて,単一スパインのアクチン構築を調べる手法を開発した。これにより,スパインのアクチン繊維が二つ のプールに分かれて存在すること,アクチン繊維の安定性が大きなスパインの方が大きいことなどが明らかとなってきた。
【追加情報】(調査月2007年8月、記入月2008年6月)
発表論文
関係論文
塩田清二(昭和大学医学部・第一解剖学)
大滝博和(昭和大学医学部・第二解剖学)
土肥謙二(昭和大学医学部・救急医学科)
岡田泰伸
虚血性神経細胞死は炎症性サイトカイン,グルタミン酸,アラキドン酸,接着分子,フリーラジカルや一酸 化窒素(NO)などの様々な因子が関与する。しかし,その詳細な機構については依然不明である。近年,細胞容積調節の異常がアポトーシス性細胞死誘導に先 立って出現し,細胞容積調節の破綻を抑制すると細胞死誘導が抑制されることが報告された(Maeno et al. 2000)。また,これまでに本共同研究により前脳虚血再潅流モデルにより誘導される虚血性神経細胞死誘導時の海馬CA1領域の神経細胞にも細胞容積の破 綻によるアポトーシス性神経細胞死が認められることを明らかにした。本年度は脳虚血神経細胞死に対しアニオンチャネルブロッカーが抑制作用を有するかどう かを機能形態学的に明らかにした。
前脳虚血モデルは,両総頚動脈を12分間血管クリップにより閉塞し,その除去により血流の再潅流をする両総頚動脈閉塞再灌流モデルを用いた (Matsunaga et al. 2003 Neurosci Res)。脳虚血直前(静脈内)および脳虚血後1,2,3日(腹腔内)にアニオンチャネルブロッカーであるDIDSおよびプロテインチロシンキナーゼ阻害薬であるgenisteinおよびvehicleを投与し,脳虚血後の神経細胞死を比較した。
脳虚血後4日目における神経細胞死をトルイジンブルー(正常細胞の計測)とFluoro-Jade B染色(死細胞の計測)により比較したところ,DIDSとgenisteinの投与群はトルイジンブルーによる形態学的正常な細胞数の低下,および Fluoro-Jade B染色による死細胞の増加を抑制した。この結果はアニオンチャネルブロッカーが神経細胞死抑制作用を示すことを示唆した。さらに,脳虚血後の海馬を細胞分 画し,ミトコンドリア画分と細胞質画分におけるチトクロームcの発現をイムノブロット法で調べたところ,DIDSおよびgenistein投与群はチトク ロームcの細胞質画分における出現が抑制されており,本細胞死の抑制がアポトーシス性神経細胞死の抑制に起因している可能性を示唆した。
【追加情報】(調査月2007年8月、記入月2008年6月)
発表論文
関係論文
酒井哲郎(琉球大学医学部形態機能医科学講座生理学第2分野)
岡田泰伸
本研究は,膜電位感受性色素を用いた細胞膜電位活動の光学測定法とCaイメージングを組み合わせた複合 的光学測定システムの開発を目指した共同研究プロジェクトである。本年度はその第二段階として,虚血のモデルとしての低浸透圧条件下における心房筋の電気 的活動の変化を調べる実験をおこなった。
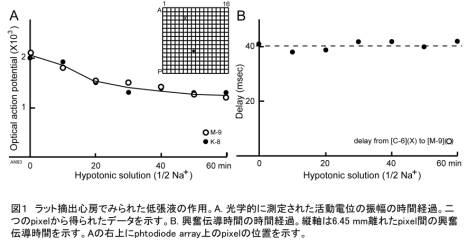
実験にはラットから摘出した右心房標本を平面的に展開した標本を用いた。この標本をメロシアニン・ロー ダニン系膜電位感受性色素NK2761で染色し,光学顕微鏡のステージにマウントした。強化・安定化した光源から波長700 nmの準単色光を標本に照射し,対物鏡と写真用接眼鏡により作られた実像面に16×16素子のフォトダイオードアレイを置き,膜電位変化を標本の最大 256ヶ所から光学的に同時記録をおこなった。筋収縮にともなう光学的アーチファクトを抑制するため,筋収縮抑制剤としてサイトカラシンD (40 mm) を作用させ,室温で測定をおこなった。
ラット心房標本をリンゲル液のNa+濃度を半分にした低張液中に置くと,図1A に示すように光学的に測定された活動電位(optical action potential)の振幅が徐々に小さくなった。その時間経過はゆっくりしており,約50-60分後にほぼ30%減少して一定値となった。しかしなが ら,図1Bに示すように,興奮伝導速度を示す離れたpixel間の活動電位の立ち上がりのタイミングの遅れ(delay) には大きな変化は見られなかった。
ここで見られた現象の発現の機序はまだ不明であるが,活動電位の振幅の低下はゆっくりとした時間経過で発生しており,活動電位の立ち上がり相のNa-spikeへの影響とは異なる機序が示唆される。また,低張ストレスに際しては細胞内Ca2+の動員が知られているが,細胞内Ca2+に感受性をもつgap junctionのconductanceの影響を受けると思われる興奮伝導速度が大きな変化を示さない点も興味深い結果といえる。今後さらに実験を加えてその実体を追跡する必要がある。
【追加情報】(調査月2007年8月、記入月2008年6月)
関係論文
高岸 芳子(名古屋大学・環境医学研究所),宮田 麻理子
ミオシンVaは神経細胞に多く発現し,アクチンフィラメントに沿ってオルガネラを輸送するモーター蛋白 である。自然発症mutantマウスであるdilute-neurological mouse (dn/dn)はミオシンVa蛋白のtail部分の14アミノ酸が欠損しているマウスであり,このマウスは生後直後から1か月程度まで,極めて激しい運動 失調,小脳失調と強直性痙攣発作を示すが,それ以降は,これらの症状が劇的に回復する特徴をもっている。我々は,これらの表現型の分子基盤,シナプス基盤 を明らかにする目的で,小脳プルキンエ細胞に着目し,解剖学的,電気生理学的,行動学的解析を行った。基本的なシナプス伝達特性に異常はなかったが,生後 1か月以内では小脳学習の一種である瞬目反射学習が獲得不能であり,またその分子基盤とされる小脳長期抑圧現象(LTD)も欠損していた。症状が回復した 生後2か月のマウスでは上記小脳学習とLTDはともに回復していた。解剖学的には,小脳プルキンエ細胞のスパインの粗面小胞体(ER)は生後1ヶ月までは スパイン内に入っておらず,樹状突起に留まるが,生後1か月以降になると,野生体と同様にスパイン内に存在することが明らかになった。小脳LTDの誘発に はプルキンエ細胞スパイン内のERからの細胞内カルシウム放出が必要とされており,本結果は,スパイン内にERが回復することによって,LTD,小脳学習 が回復すると考えられた。今後,ERの動態とミオシンVaの機能関連ついて更なる研究が必要である。
【追加情報】(調査月2007年8月、記入月2008年6月)
発表論文
饗場 篤(神戸大学・大学院医学系研究科),宮田 麻理子
代謝型グルタミン酸受容体 (mGluR) のうち,mGluR1はmGluR5と共にグループIを形成し,三量体G蛋白質Gqと共役しプロテインキナーゼC (PKC) 活性や細胞内カルシウム濃度を調節している。本研究では,mGluR1サブタイプのうち,mGluR1aが持つ長い細胞内C末端ドメインを欠く mGluR1bサブタイプのみを発現するマウス小脳皮質でのシナプス伝達,シナプス可塑性等を測定した。結果,通常のシナプス伝達には異常がなかったが, 小脳長期抑圧現象(LTD)はmGluR1aレスキューマウスでは回復するものの,mGluR1bレスキューマウスでは欠損していており,C末端ドメイン -homer蛋白以下のシグナルがLTDには必須であることが明らかになった。今後,脳の他の部位についても解析を進める予定でいる。
高田昌彦(東京都神経科学総合研究所)
泰羅雅登(日本大学大学院総合科学研究科)
南部 篤
大脳基底核の機能を考える上で,大脳皮質との相互連絡,さらには大脳皮質間の線維連絡を知ることが重要 である。その際,特に経シナプス的に情報がどのように伝達されるのかに注目することが重要であるが,従来の方法では捉えることができなかった。狂犬病ウイ ルスがシナプスを超えて逆行性に感染することを利用して,これをトレーサーとして用い,大脳皮質間での線維連絡の解析を行った。なお,ウイルスの注入は東 京都神経科学総合研究所の感染実験室で行った。
一次運動野上肢領域に狂犬病ウイルスを注入し,経時的に観察した。注入後2日では高次運動野の尾側部, 視床運動核などがラベルされた。3日目では,高次運動野の吻側部や弓状溝前方の8野などに広がった。また,淡蒼球内節や小脳核にもラベルが見つかった。4 日目では,前頭前野全域,被殻,小脳皮質までラベルが広がった。また,一次運動野上肢領域と下肢領域に狂犬病ウイルスを注入し,それぞれに投射する領域を 調べたところ,上肢領域注入例では46野腹側部を中心に前頭前野に多くラベルが見られたが,下肢領域注入例ではあまり観察できなかった。これは,46野腹 側部が上肢運動の制御に重要な役割を果たすことを示唆している。
【追加情報】(調査月2007年8月、記入月2008年6月)
関係論文
稲瀬正彦,中陦克己(近畿大学医学部)
森 大志(山口大学農学部)
南部 篤
脊椎動物の歩行運動において,姿勢および四肢のリズム運動を制御する基本的な神経機構は脳幹・脊髄に分 散的に配置される。一方ヒトが歩行の速度を増加させる場合には,遊脚相時間を一定に保つ傍ら着地相時間を短縮させ,結果として歩行周期頻度を増加させる。 この様な速度増加に対する歩行パラメータの対応関係は,イヌやネコ或いはサルといった四足動物においても同様に認められる。これらのことは二足歩行制御に おいて,四足動物と相同な脳幹・脊髄の基本的神経機構がヒトにおいても存在することを推察させる。
流れベルト上で二足歩行するサルは,歩行速度の増加に対して歩行パラメータをヒトと同様に変化させる。 一方,大脳皮質運動領野は主として脳幹・脊髄に出力する。本課題では二足歩行制御および四足歩行制御との間における共通の神経基盤の存在を検証することを 目的とした。そのために流れベルト上で四足歩行および二足歩行を交互に遂行することを学習したサルの一次運動野・下肢領域から神経細胞活動を導出記録し て,四足歩行時における活動様式を二足歩行時のものと比較した。
四足歩行において,一次運動野・下肢領域から記録された神経細胞の殆んどは,歩行周期に伴って相動的に それらの活動を修飾した。相動性に活動した神経細胞は1~2 峰性の修飾様式を示した。これらの修飾活動は主に歩行周期における着地相に認められたものが多く,遊脚相にみられた活動は比較的少なかった。四足歩行にお いて相動性の活動様式を示した神経細胞の殆んどは,二足歩行においても同様に相動性の活動様式を示した。歩行周期中における発射頻度のピークの時期は,四 足歩行時のものとほぼ一致した。二足歩行時における平均発射頻度ならびに一歩行周期中における発射頻度のピーク値は,四足歩行時のそれらに比べて有意に高 値を示した。四足歩行時或いは二足歩行時において選択的に活動する神経細胞も少数ながら記録された。
以上の結果は四足歩行時および二足歩行時において,歩行周期に同期した一次運動野からの出力を受ける共 通の皮質下神経機構の存在を示唆しており,サルの一次運動野が,それらの皮質下神経機構に対する四足歩行時の出力様式を修飾し,さらには新たな皮質からの 出力を動員することによって二足歩行運動を発動・制御することを示唆する。
【追加情報】(調査月2007年8月、記入月2008年6月)
発表論文
木下 専(京都大学大学院医学研究科)
萩原 明(京都大学大学院医学研究科)
重本 隆一(生理学研究所)
セプチンは酵母からヒトまで保存された重合性GTP結合蛋白質であり,細胞膜の裏打ちに集積してスカ フォールドを形成する。出芽酵母のセプチンは分裂溝の細胞膜直下で環状構造体(セプチンリング)を形成し,これが分裂関連蛋白質の足場として,あるいは膜 蛋白質などの拡散障壁として,細胞分裂に必須であることが確立している。一方,多細胞動物のセプチンは分裂後のニューロンやグリアにも多く発現している が,その機能は不明である。我々は,ニューロンやグリアの形態形成やシナプス関連蛋白質の局在におけるセプチンの重要性を検証するため,セプチン及びセプ チンと生化学的ないし機能的に相互作用する蛋白質の局在をマウス脳切片や培養系を用いて解析している。セプチン複合体は密な高次集合体を形成するため,従 来の免疫電子顕微鏡法では十分な検出ができない可能性もある。そこで本共同研究では,蛋白質が密に集積している場合でも高感度の検出能を有する凍結割断レ プリカ免疫標識法を,小脳や海馬に適用して局在解析を試みた。
従来の免疫電子顕微鏡法を用いた研究で,セプチンSept5がGABAニューロンの前シナプス末端に集 積することを報告したが,凍結割断レプリカ免疫標識法では海馬のシナプス後肥厚部 (PSD) においてPSD-95と共在していることが今回新たにわかった。すなわち,セプチンの一部はPSDにおいて神経伝達物質受容体などの後シナプス蛋白質のス カフォールドとして機能する可能性が示唆された。しかしながら,小脳では従来の免疫電子顕微鏡で観察されていたリング状のセプチン (Sept4) クラスターが,レプリカ標識では散在性のシグナルとして検出され,2種類の方法で観察される局在様式に相関性が乏しかった。したがって,ニューロンやグリ アにおけるセプチンの分布様式を明らかにしていくためには,さらなる検証が必要である。
【追加情報】(調査月2007年8月、記入月2008年6月)
関係論文
横井峰人(京都大学医学研究科・先端領域融合医学研究機構・分子神経遺伝学グループ)
佐野裕美(京都大学医学研究科・先端領域融合医学研究機構・分子神経遺伝学グループ)
線条体投射神経細胞は,黒質,淡蒼球へ投射するのみならず,外側視床下部領域への投射が存在することが 示唆されている。当研究では,線条体複合体(被殻−尾状核,側坐核,嗅結節)の投射神経細胞に選択的にGFPあるいはsynaptophluorinを発 現するトランスジェニックマウスを作成し,そのマウスを用いて線条体投射神経回路の投射領域を可視化することを試みた。その結果,線条体投射神経細胞は, 外側視床下部領域においてある限られた領域へ投射することが明らかとなった。さらに,この投射領域の性質を各種化学マーカー,トーレーサーにより解析をす すめている。
また,線条体投射神経細胞に極めて豊富に存在するphosphodiesterase PDE10Aの抗体を作成し,免疫電顕にて細胞内局在を検討した。その結果,PDE10Aは細胞質に広く分布するのではなく,また,膜組織上にはないもの の,樹状突起棘内においてシナプス近傍に多く分布するという特徴ある細胞内局在を示した。このことは,PDE10Aが,細胞内情報伝達の重要な担い手の一 つであるcAMPの樹状突起棘内におけるシナプス近傍からの拡散を防ぎ,cAMPの細胞内局所濃度勾配に寄与する可能性が示唆された。
【追加情報】(調査月2007年8月、記入月2008年6月)
発表論文
関係論文
伊藤 功(九州大学大学院理学研究院)
我々は成獣マウス海馬シナプスのNMDA受容体応答およびNMDA受容体サブユニットのシナプス分布を 解析することにより,海馬神経回路は左右の海馬および錐体細胞の上下に関して非対称であることを発見した。本共同研究ではこの発見をさらに発展させ,脳の 左右差の形成時期とこれに関与する遺伝子の解明。脳の左右差の行動学的意義の解明。および左右非対称な神経回路の形成に関与する細胞外シグナルの解明,を 目的として総合的な検討を行った。本年度は,内臓に左右の逆位を示すivマウスの海馬を用いた解析を行い,その結果,ivマウスでは左右の非対称性が消失 し,神経細胞の上下の非対称性だけが正常であることが明かになった。我々の結果は,内臓等の内胚葉由来の組織と,脳のような外胚葉由来の組織では,左右差 の形成機構が異なっていることを示唆している。
【追加情報】(調査月2007年8月、記入月2008年6月)
発表論文
柳川右千夫,柿崎利和,白尾智明,齊藤康彦(群馬大学・大学院医学系研究科)
川口泰雄,平林真澄(生理学研究所)
大脳皮質におけるリズミックな神経活動,神経が同期して活動する現象,シナプス可塑性の形成には,抑制 性ニューロンが行う神経情報処理が重要な役割を果たしていると考えられている。しかしながら,抑制性ニューロンは,多様な形態をもち,in vitroおよびin vivoで正確に同定することは困難である。そこで,抑制性ニューロンを蛍光分子で標識したトランスジェニックラットを作成し,解析を行った。抑制性 ニューロンに特異的発現する小胞型GABAトランスポーター(VGAT)遺伝子に蛍光蛋白分子をコードするVenus遺伝子を挿入したコンストラクト (VGAT-Venus) を作成した。このコンストラクトをラット受精卵に導入することにより,トランスジェニックラット(VGAT-Venusラット)を作成した。2系統の VGAT-Venusラットを繁殖させ,解析に用いた。VGAT-Venusラット大脳皮質GABAニューロン全体に対するVenus発現細胞の割合につ いて,Venusの蛍光と抗GABA抗体による2重染色を用いて検討した。その結果,2系統ともに殆どすべてのGABAニューロンでVenus分子の発現 を観察した。また,大脳皮質のVenus陽性細胞からホールセル記録を行った結果,GABAニューロンに特徴的なfast-spiking等の発火パター ンを観察した。以上の結果は,大脳皮質のGABAニューロンの組織学的解析や電気生理学的解析にVGAT-Venusラットが有用であることを示唆する。 また,Venus遺伝子の代わりにカルシウムインディケーター遺伝子などをVGAT遺伝子に挿入することで,抑制性ニューロンの機能解析用遺伝子改変動物 の作成が可能となった。
大脳皮質抑制性ニューロンの形態学的および電気生理学的特性を理解するには,他の領域における抑制性 ニューロンに関する知見が重要である。そこで,VGAT-Venusラット前庭神経核と舌下神経前位核に存在する抑制性ニューロンの組織学的解析を行っ た。また,抑制性ニューロンをEGFPで標識したGAD67-GFPマウスを利用して,扁桃体抑制性ニューロンへのドーパミンの作用など,様々な抑制性 ニューロンの機能や形態について明らかにした。
【追加情報】(調査月2007年8月、記入月2008年6月)
発表論文
根東 覚(東京医科歯科大学・大学院医歯学総合研究科・細胞生物学分野)
川口泰雄(生理学研究所)
藤田一郎(大阪大学大学院・生命機能研究科)
狩野方伸(金沢大学・大学院医学系研究科)
前頭皮質は大脳基底核の入力部である線条体に強い出力繊維を送り,線条体の神経回路の活動を調節してい る。たとえば,線条体のGABA作動性投射細胞は特徴的な2層性の膜電位遷移状態をとることが知られているが,このリズムは皮質の細胞の律動的活動と関係 していることが分かってきている。一方皮質-線条体路をつかさどる細胞は,大脳皮質の主に第5層にある錐体細胞であることが既に知られているが,皮質内の 回路に於いてその律動的な活動がどのように調節されているかはまだ未解明である。その機構の一つとして錐体細胞間に形成される反回性の興奮回路が考えられ るが,それだけでは説明がつけられない。皮質内回路には興奮性の投射細胞以外に多種の介在性GABA細胞が存在し,回路の興奮性を微妙に調節している。今 回私は,これらの介在細胞の中で,特に錐体細胞の発火の調節に重要と考えられる高頻度発火型介在神経細胞(FS細胞)と線条体投射錐体細胞の神経結合を, ラット脳から調整した急性スライス標本を用いて逆行性標識と2細胞同時記録を行うことにより電気生理学と形態学的な解析から調べた。錐体細胞の細胞体から パッチクランプ法を用いて膜電位固定によりシナプス電流を計測すると,その振幅の大きさから結合の強さにはペア間にばらつきがあることが分かった。また, シナプス前細胞の連続的な2回の発火に対してシナプス電流の大きさはいずれも1回目に対して2回目は減弱を示し,その比率はほぼ一定していた。
記録を行った細胞を細胞内染色法により可視化して3次元再構築を行うと,FS細胞は,細胞体へのシナプ ス結合比率から所謂バスケット細胞と考えられたが,シナプス結合部位は錐体細胞の細胞体周辺だけでなく,比較的遠位の樹状突起にもあることが分かった。た だ,樹状突起上への結合は主として基底樹状突起に見られ,尖頭樹状突起には少なかった。シナプス電流の振幅とシナプス結合の細胞体からの距離について調べ てみると,結合の数とは相関が見られなかったが,最も近い距離とは正の相関が見られた。FS細胞の軸索と錐体細胞の樹状突起の近接点を調べ,シナプス結合 部位との関係を調べると近接点のうち10%程度においてシナプスが形成されており,近接点の分布とシナプス結合の分布にほとんどのペアにおいて優位な相関 関係が見られた。
【追加情報】(調査月2007年8月、記入月2008年6月)
関係論文
宇賀 貴紀(順天堂大学医学部)
神作 憲司(生理学研究所大脳皮質機能研究系)
立体視とは両眼視差,すなわち物体が両眼に落とす網膜像の位置のズレから生じる奥行き知覚のことであ る。サル大脳皮質視覚野では,背側経路(空間視経路)で絶対視差(1つの物体の注視点に対する両眼視差)の情報処理が,腹側経路(物体視経路)で相対視差 (2つの物体の絶対視差の差)の情報処理が行われていると考えられている。本研究では,これらサルでの知見を基に,人でも絶対視差・相対視差情報の並列処 理が行われているのか,それぞれの情報処理がどの脳部位で行われているのかを検証する。
本研究では,絶対視差・相対視差それぞれを弁別中に,脳内磁気刺激を用いて特定部位の脳活動を一時的に 休止させ,絶対視差・相対視差に特異的な情報処理過程の欠落が見られるか検討する。絶対視差検出課題では,被験者は1つの物体が注視点の手前にあるのか, 奥にあるのかを答える。相対視差検出課題では,被験者は2つの物体のどちらが手前にあり,どちらが奥にあるのか,その相対関係を答える。被験者がこれらの 課題を遂行中に,大脳皮質MT野を磁気刺激する。サルのMT野は絶対視差のみを処理していることがわかっている。もし人でも同様であれば,絶対視差の検出 は障害されるが,相対視差の検出は障害されないと予想される。現在プログラムの作成中であり,まだデータを取得してない。今後は予備実験を施行後,本実験 に移る予定である。
大木紫(杏林大学医学部統合生理学教室),小川潤(杏林大学医学部整形外科学教室)
伊佐正,西村幸男(自然科学研究機構生理学研究所・科学技術振興機構研究員(生理学研究所))
脳からの運動指令を運動ニューロンに伝える皮質脊髄路は,従来運動ニューロンに単シナプス性結合する直 接的経路のみが重要視されてきた。しかし近年,脊髄内中継ニューロンを介する間接的経路が同等以上に機能することが明らかになった。特に脊髄で直接的経路 が障害を受けた場合,間接的経路を用いて機能回復が起こる可能性が示唆されている。本研究では間接的経路の神経回路の性質について,麻酔下のサルにおいて 電気生理学的手法を用いて解析を進めた。
行ったのは,ネコで間接的経路の中継ニューロンとして知られる,第3-4頸髄の脊髄固有ニューロン (PN) からの直接記録である。麻酔下サルの第3-4頸髄に記録電極を刺入し,177個の脊髄介在ニューロンの細胞内,または細胞外の電位変化を記録した。記録中 に皮質脊髄路を刺激して入力を観察,さらに下部頸髄を電気刺激して運動ニューロンへの投射の有無を確認した。細胞外記録した介在ニューロンのうち,皮質脊 髄路の3連発刺激により活動する細胞は21%しか存在しなかったが,ストリキニンで抑制性入力を遮断するとほとんどの細胞(83%)で活動が観察されるよ うになった。運動ニューロンへ投射するPNではストリキニン投与により初めて活動が確認できるものが多く,投与前の細胞内記録により単シナプス性興奮性入 力と直後に2シナプス性抑制性入力が誘発されることが確認された。次にPNの運動ニューロン以外への投射を見るため,延髄の外側網様核の電気刺激を行っ た。ネコのPNはほとんど(84%)が外側網様核にも投射することが知られているが,サルのPNでは30%の細胞しか投射が確認されなかった。
今回,皮質から運動ニューロンへの入力を中継する脊髄ニューロンが,霊長類では始めて直接的に確認され た。直接的皮質脊髄路入力が発達した霊長類ではこの経路が退化したという報告が以前なされたが,これは抑制性入力が興奮性入力を打ち消していること,外側 網様核への投射細胞が霊長類では少ないことを考慮しなかったためと考えられた。ストリキニンを投与しない麻酔下の動物では,PNを介した間接的経路はほと んど活動しない。しかし覚醒運動時の動物では活動する可能性もあり,確認する必要がある。また直接的経路障害時にはPNを介した間接経路が活性化されるこ とを我々がすでに報告しており,この経路の機能回復への関与も更に検討する必要がある。
【追加情報】(調査月2007年8月、記入月2008年6月)
発表論文
大石高生(京都大学霊長類研究所),肥後範行,村田弓(産業技術総合研究所)
伊佐正(自然科学研究機構生理学研究所)
ヒトをはじめとする霊長類は指を独立に動かすことにより,物体の操作などの器用な運動を行うことができ る。系統発生的に見て霊長類で非常に発達している皮質脊髄路がその神経基盤の一つである。Kuypersらの古典的な研究では,皮質脊髄路を延髄レベルで 切断すると指の器用な運動が永久に消滅した(Lawrence and Kuypers, 1968)。しかし,延髄での切断は他の運動経路の破壊も含むものであり,頚髄レベル(C4/5) で皮質脊髄路を切断すると,運動機能が回復することが近年の伊佐らによって示された(Sasaki et al., 2004)。われわれは神経回路の構造的変化による代償が運動機能回復の基盤になっているという仮説を検討するため,構造変化のマーカー分子の一つである GAP-43の遺伝子発現とタンパク発現を皮質脊髄路損傷ザルで組織学的に検討した。
解析に用いたのは頚髄レベルで皮質脊髄路を切断した後,訓練により精密把握ができるように回復した二頭 のニホンザルで,一頭は運動機能が回復して3.5ヶ月経った(損傷後4ヶ月)個体(C),もう一頭は運動機能が回復して1ヶ月経った(損傷後3ヶ月)個体 (G)であった。
大脳皮質運動関連領野でGAP-43に対するin situハイブリダイゼーションを行ったところ,個体Gでは皮質脊髄路切断部と反対側の一次運動野の上肢領域,運動前野腹側部,背側部で同側に対して GAP-43遺伝子の発現が上昇していた。発現上昇は深層でも浅層でも起こっていた。補足運動野では発現上昇は見られなかった。運動機能回復後時間が経っ た個体Cではいずれの運動関連領野でも切断部の反対側同側を問わず遺伝子発現の上昇が見られなかった。
脊髄でGAP-43に対する免疫組織化学を行ったところ,個体Gでは,損傷部位に近い髄節の外側皮質脊髄路,側索の前角近傍部,脊髄小脳路で,破壊側の方が反対側よりもGAP-43を強く発現する構造が多く見られた。個体Cでも同様の傾向が見られた。
これらのことから,頚髄レベルの皮質脊髄路切断後の運動機能回復時には,少なくとも大脳皮質の複数の運動関連領野および脊髄で軸索伸長等の神経回路の構造変化が起こっていること,大脳皮質での変化は運動機能回復が完成して一定時間が経過すると停止することが推測される。
【追加情報】(調査月2007年8月、記入月2008年6月)
発表論文
小林康(大阪大学大学院生命機能研究科)
伊佐正(自然科学研究機構生理学研究所)
【研究計画】 中脳脚橋被蓋核(PPTN) は脳幹のもっとも大きなアセチルコリン性細胞の核であり,古くから睡眠覚醒の調節,運動制御,注意や学習と関係が深いと考えられてきた。PPTNは眼球運 動中枢や脳幹や脊髄の運動中枢に投射しており,上行性には報酬に基づく学習(強化学習)に関係していると考えられている大脳基底核の黒質緻密部に強力に投 射しており,さらに,PPTNが運動制御,強化学習や動機付けと関係していることを示す破壊実験や生理実験が多く示されてきた。さらに,PPTNが外側膝 状体,青斑核,縫線核と結合しているということを考えあわせると,PPTNを中心とした神経回路で注意のシステム,動機付け,運動系,報酬系が密接に関係 していると思われる。最近申請者はサルを用いた行動生理実験でPPTNがサッカード制御(運動準備,運動実行,誤差信号),報酬情報の処理,さらに報酬量 制御による動機付けの度合いの調節と関係していることを明らかにしている (Kobayashi et al, 2002, 2004)。本研究では,腕の到達運動やサッカードの制御,学習に脚橋被蓋核が果たす役割を解析することを目的とした。ところで,中脳黒質,腹側被蓋野 ドーパミン細胞(DAcell)は報酬で条件付けされたcueや報酬に対してphasicなバースト応答をすることによって大脳基底核などに報酬予測誤差 を送り,強化学習におけるシナプス可塑性を制御していると考えられている。DAcellにおいて報酬予測誤差がどうやって計算されるかということは強化学 習機構を解明する上で最も重要な問題の一つであると考えられている。PPTNはDAcellに強力に投射していることからPPTNからの興奮性入力 が,DAcellにおける報酬予測誤差信号の生成に重要な役割を果たしていることが示唆される。本研究でその問題にもアプローチした。
【研究結果】 ところで,特に報酬に基づく強化学習においては大脳皮質や大 脳基底核等では学習される行動に特化したニューロン活動が表現され,むしろ中脳ドーパミン細胞(DAcell)やPPTNでは行う行動には無関係に報酬の 価値が表現されている可能性がある。行う行動と無関係な報酬価値のみに関係するニューロン活動を記録するため,頭部に固定具を装着した覚醒ニホンザルをモ ンキーチェアに座らせ,頭部を固定し,コンピューターディスプレイ上に表示された光点の指示に従ってモンキーチェアに設置したボタン押し(上肢による)と サッカード運動(眼球運動)を行わせた。上肢運動,眼球運動といった異なる運動の学習に関係するニューロン活動を記録する前に本研究ではまず,サルに手が かり刺激で報酬量を予測させるような視覚誘導性サッカードを行わせ,その時のPPTNのニューロン活動を記録した。報酬予測サッカード課題中のサル PPTNニューロンの活動を記録すると,報酬予測の度合いによって大きさが変わるcue呈示から短潜時で起きる持続的応答と,報酬前から起こる持続的活動 の減少,報酬予測の度合いに無関係に常に一定の大きさの報酬反応等の結果が得られた。以上の結果はPPTNニューロンがサッカード運動にかかわらず報酬予 測,報酬に関係した活動を示すことを表している。また,PPTNからの興奮性入力が,DAcellにおける報酬予測誤差信号の生成に重要な役割を果たして いることを示唆している。今後,サルが上肢によるボタン押しや上肢の到達運動とサッカード運動を行っているときにニューロン活動を記録する予定である。
【研究業績】
Y.Kobayashi, et al. REWARD-PREDICTING ACTIVITY OF PEDUNCULOPONTINE TEGMENTAL NUCLEUS NEURONS DURING VISUALLY GUIDED SACCADE TASKS Soc.for Neurosci.Abstr.35 890.5, 2005
M.Watanabe, et al. INFLUENCE OF INSTRUCTION AND STIMULUS POSITION OF THE PREVIOUS TRIAL ON
REACTION TIMES OF PROSACCADES AND ANTISACCADES IN HUMANS Soc.for Neurosci.Abstr. 35 859.12, 2005
Yasushi Kobayashi, et al. Reward predicting activity of Pedunculopontine Tegmental nucleus neurons during visually guided saccade tasks Neurosci. Res Suppl.28 O3F-06
Kenichi Okada, et al. Visual and oculomotor functions of monkey pedunculopontine tegmental nucleus Neurosci. Res Suppl.28 P2-152
小林悠一他登上線維入力が小脳プルキンエ細胞の活動に与える影響電子情報通信学会技術研究報告NC2005-39 pp.13-19, 2005
【追加情報】(調査月2007年8月、記入月2008年6月)
関係論文
林 弘之(神奈川歯科大学)
鍋倉 淳一,和気 弘明,堀部 尚子(生理学研究所)
成熟後の神経再生期には伝達物質受容体や細胞内環境などにおいて一旦幼若期の特徴が再出現する。そこ で,抑制性神経回路が障害脳において未熟期の特徴を再獲得する可能性について検討するために,本共同研究を行った。具体的には,未熟期において,三叉神経 運動核へのGABAおよびグリシン作動性終末が一旦余剰に入力し,発達とともに次第にGABA・グリシン終末が減少していくこととの本研究の共同研究者で ある林らの報告を受けて,三叉神経運動細胞の軸索傷害後の軸索再生期に,同核におけるGABAおよびグリシン入力の変化を免疫染色および電気生理学的手法 を用いて検討した。まず,三叉神経運動細胞の軸索の同定,および障害細胞自体の核内分布を調べるため,軸索傷害後,中枢側に親脂質性蛍光色素(DiI)を 投与し,数日後に,同運動核内におけるDiI陽性細胞の分布を検討した。次に,同部位におけるGABA/グリシン入力の障害後における変化をGADおよび グリシンに対する抗体を用いて検討した。また,終末自体の変化をsynaptophysin抗体を用いて検討した。軸索傷害,1週間後で,三叉運動神経核 外側部におけるGABA染色性の強い部分が観察された。しかしながら,その他の部位ではGABAおよびグリシン染色性の著明な変化は観察されなかった。ま た,synaptophysion抗体の染色性は,やや増加傾向にあるものの著明な変化は確認できなかった。
電気生理学的には傷害1週間後にはGABA応答とグリシン応答の比として観察すると,GABA優位であり,その後ややグリシン優位に変化する可能性が示唆される結果が得られた。しかし,いずれも著明な変化として定量的に解析可能か今後更に検討する必要がある。
島津 和弘(国立長寿医療センター)
鍋倉 淳一,溝口義人(生理学研究所)
幼若期においてはGABAによって脳の多くの部位では神経細胞成熟や神経回路成熟が調節されていること が近年,報告されている。GABAの応答は主な細胞内Cl濃度調節因子であるKCC2(神経特異的Clくみ出し分子)とNKCC1(Clくみ入れ分子)に 機能発現により制御されている。幼若期には,KCC2の機能的未発達に起因する細胞内高Cl濃度のため,GABAは脱分極作用を示し,細胞内Ca濃度上昇 などを引き起こす。しかし,このKCC2の発現調節に関しては殆どわかっていない。一方,海馬歯状回では成熟期においても神経細胞新生が起こる。また,同 部位ではGABA陽性の顆粒細胞が一定割合で存在する。そこで,GABA陽性細胞と新生未熟神経細胞,KCC2発現,およびこれらを制御するニューロトロ フィンの関連の検討を試みた。Brdu陽性新生細胞におけるGADの有無,およびKCC2発現を検討した。Brdu陽性細胞とGAD陽性細胞は一定割合で 一致したが,明確な関連についてはBrdu染色法によるGAD染色性の脆弱化のため,今後の共染色技術の改良が必要である。また,培養海馬細胞におい て,BDNFが時間単位でKCC2を脱燐酸化し,その後蛋白自体の発現を著明に低下させることが判明した。新生神経細胞に対するBDNFのGABA発現お よびKCC2に対する作用を調べるため,現在スライス培養法による検討を試みている。
平田雅人,兼松隆(九州大学)
鍋倉淳一,溝口義人(生理学研究所)
PRIPとは,我々が発見し遺伝子クローニングを行った,新しいイノシトール1,4,5,-三リン酸結合性タンパク質である。この分子は,PLC-d1と高い相同性を有するが,PLC 酵素活性を持たない。そこで,PLC-related but catalytically inactive protein (PRIP)と名付けた。最近,PRIP-1 KOマウスを解析して,この分子が抑制性神経伝達の一角をなすGABAのシグナル伝達に重要な分子であることを突き止めた。そこで,自然科学研究機構生理学研究所 鍋倉研究室の協力を得て「GABA シグナリングにおける新規分子PRIPの役割解明研究」を遂行している。
我々は,PRIP-DKOマウス(PRIP-1 & -2ダブルノックアウトマウス)を作製し,その初代神経培養細胞を用いて,BDNF(脳由来神経栄養因子)刺激で誘導されるGABAトランスミッションの 反応性の違いを,野生型と比較検討した。すると,PRIP-DKOマウス初代神経培養細胞では,GABA反応が野生型と全く逆になることを突き止めた。そ こで,BDNFで誘導されるGABAシグナリングのどの過程にPRIP分子が関与するか,その分子基盤の解明を目指して実験を行った。その成果を以下に列 挙する。
1,PRIP分子は,プロテインホスファターゼ (PP1・PP2A) に結合してGABAA受容体のbサブユニットの脱リン酸化過程を修飾する。
2,PRIP分子が欠損してGABAA受容体bサブユニットの脱リン酸化反応が阻害されると,受容体のエンドサイトーシスが抑制された。
3,PRIP分子は,GABAA受容体のbサブユニットに結合するが,その結合を阻害するペプチドを用いると,野生型神経細胞においてみられたBDNFで誘導されるGABA-induced Cl- 電流の減弱効果を完全に阻害することが出来た。
以上から以下のことが明らかとなった。
PRIP分子は,GABAA受容体のbサブユニットに結合しプロテインホスファターゼを受容体にリクルートすることで,受容体の脱リン酸化過程を調節する。その結果,PRIPにより脱リン酸化された受容体のエンドサイトーシスが促進される。
この成果を右論文に発表した。Kanematsu et. al., (2006) J. Biol. Chem., in press.
いくつかのグループが,GABAA受容体のエンドサイトーシス過程はクラスリン/AP2複合体によって誘導されると報告している。現在我々は,クラスリン/AP2複合体により誘導される受容体のエンドサイトーシスにPRIP分子がどの様に関わるかをさらに詳しく検討している。
【追加情報】(調査月2007年8月、記入月2008年6月)
発表論文
関係論文
美馬達哉(京都大学大学院医学研究科)
逵本徹
ヒトの脳波活動と上肢の筋電図活動の間には相関が認められる。この相関関係は大脳皮質運動野から筋肉へ の運動制御過程を反映している可能性がある。しかし,一次運動野が近隣の運動関連領野と関係しながら筋収縮をコントロールしていることは一見当然とも考え られるにもかかわらず,その神経機構は十分には解明されていない。我々は,動物実験を行うことによってこの仕組みを神経回路レベルで解明できる可能性があ ると考え,サルの大脳皮質フィールド電位と上肢筋電図活動の記録及び解析を行った。その結果,筋電図活動と相関を示す大脳皮質の部位を特定することができ た。さらに運動の実行中にこの相関関係がどのように変化するかを明らかにして,運動制御の動的な神経機構を研究する予定である。
福井豊(帯広畜産大学・畜産学部・家畜増殖学研究室)
保地眞一(信州大学・繊維学部・資源生物学講座)
平林真澄, 加藤 めぐみ(生理学研究所・遺伝子改変動物作製室)
【クロミンククジラ精子の中心体に由来する星状体の形成とチューブリン繊維網の成長について】
げっ歯類を除く多くの哺乳動物では,受精時に精子によって持ち込まれ中心体から星状体ができ,そこから 発達したチューブリン繊維網が雌雄前核卵子の中央部に移動させる。平成17年度では,ヒト精子中心体の機能解析に利用されるウシ卵子への異種顕微授精系に おいて,南極海産クロミンククジラ精子がもつ微小管形成中心としての能力を評価した。
方法は−20℃で保存しておいたクロミンククジラ精子およびウシ精子を融解し,5 mM dithiothreitolで5~20分間処理後,ウシ体外成熟卵子に顕微注入 (ICSI)した。ICSI後4~6時間目に微小管安定化バッファーとメタノールで卵子を固定し,免疫蛍光染色によってa-チューブリンを,DAPIによって核を染色して精子星状体の形成率を求めた。また,クジラ精子注入卵の一部は7%エタノール(5分)と2 mM 6-DMAP(4時間)の併用処理により活性化誘起し,ICSI後4時間目に星状体形成率とチューブリン繊維網の対卵子径比を調べた。
クジラ精子から星状体が形成された卵子の割合は33% (10/30) で,ウシ精子由来の星状体形成率 (33%,11/33) と同等であった。星状体形成卵のうち注入精子により活性化が起こっていたのは6例 (18%)であったのに対し,クジラでは2例 (7%) のみであった。活性化処理を行った場合,精子星状体形成率の増加(35%,11/31,対照区:26%,8/30)は僅かだったが,チューブリン繊維網のサイズは0.30から0.57に改善された。
以上の結果から,クジラ精子では中心体を起点として星状体が形成され,そこからのチューブリン繊維網の成長には卵細胞質環境が影響することが示唆された。
保地眞一,岩浪亮人(信州大学大学院・工学系研究科)
平林真澄(生理学研究所・遺伝子改変動物作製室)
マウスでは顕微操作に依存しない外来遺伝子導入法として遺伝子溶液を注入した精巣にエレクトロポレー ションをかける「精巣経由の遺伝子導入法」が報告されている。本研究では新生仔雄ラットの精巣にEGFP遺伝子を打ち込んでエレクトロポレーションをか け,生殖幹細胞ゲノムに直接EGFP遺伝子を導入することを試みた。2~3 週齢のWister系雄ラットの精巣網から,先端内径10~20mmのガラスピペットを用いて外来遺伝子溶液(濃度1 mg/ml,5.5 kb pCX-EGFP プラスミドをSal Iで直鎖化し,0.04%トリパンブルー加TEバッファーに溶解したもの)を注入した。そして精巣を接触 部直径5 mmのピンセット型電極で挟み,スクエア波形が出力できるエレクトロポレーターによって静電パルス(35~50 V, 50 msec, X方向とY方向各4回)を与えた。6週間後に精巣を摘出して蛍光実体顕微鏡下で観察したところ,EGFP タンパク質の長期発現部位のほとんどは精細管の基底膜付近で認められ,その細胞群の形態から体細胞系の細胞へ入ってしまったものと推測された。またそれら とは違う細胞群で蛍光を発した精細管も観察されたが,その割合は極めて少なかった。蛍光が認められた精細管から調製した精細胞1,526個のうち円形精子 細胞で4個 (0.3%),伸張精子細胞で10個 (0.7%) にEGFP蛍光が認められた。これらにはEGFP遺伝子が導入されていたと示唆され,たぶんその起源となる精原細胞のゲノム上にもEGFP遺伝子が乗って いたものと思われる。しかしこれらの蛍光精子細胞を顕微授精し,胚移植してみたが,産仔を得るには至らなかった。今後,この方法による外来遺伝子導入効率 の改善に向け,エレクトロポレーション条件の見直しを図る。
【追加情報】(調査月2007年8月、記入月2008年6月)
発表論文
関係論文
臼田信光,中沢綾美,谷口孝喜,厚沢季美江(藤田保健衛生大学・医学部)
金子康子,仲本準(埼玉大学・理学部)
小柳 義夫(京都大学・ウイルス研究所)
前島一博(理化学研究所)
登阪雅聡(京都大学・化学研究所)
Rasmus Schroeder(マックスプランク研究所)
永山國昭(岡崎統合バイオ)
高コントラストを特徴とする2つの電子顕微鏡法,ゼルニケ位相差法と微分干渉法(ヒルバート微分法)を 用いて応用研究を行った。高加速の300kV電顕を用いた位相差法では,氷包埋を行った細胞や組織を材料として電子染色なしで高コントラストの電顕像が得 られた。培養全載細胞観察においてはミトコンドリア等の細胞小器官と微小管等の細胞骨格が明確に認められた。血小板や精子においては細胞内部構造が立体的 に観察された。同様の観察はウィルスやバクテリアにも適用可能であった(Kaneko Y et al., J Electron Microscop 54, 79-84, 2005)。また,対象物を染色剤や蛍光剤で組織化学的に標識して,同一細胞を光学顕微鏡と位相差電子顕微鏡で観察する方法を試み,アクチンフィラメン ト,微小管の光顕-電顕相関イメージを得た。
【追加情報】(調査月2007年8月、記入月2008年6月)
発表論文
関係論文
加藤幹男(大阪府立大学・総合科学)
永山國昭,片岡正典(岡崎統合バイオ)
本研究では,生物ゲノムに存在する特徴的な塩基配列が,染色体構築と遺伝子機能制御におよぼす効果を明らかにすることを目的として,まずサテライト,ミニサテライトに代表される縦列型反復DNA配列に注目し,その高次構造特性の解析を進めている。
海水魚キチヌAcanthopagrus latusゲノムに存在する30bpを単位鎖長とするミニサテライト配列AL79は,近縁種のマダイPagrus majorには存在せず,その一方で,キチヌゲノム内には多数散在していることがわかっている。すなわち,すくなくとも,これらの種 が分岐後にAL79配列が生成し,その後の進化的に短い時間内にキチヌゲノム内で分布を広げたことを示唆する。このような現象をもたらすDNAの特徴を明 らかにするために,生化学的構造解析とともに,電子顕微鏡による構造解析を行った。AL79領域を含む組換えプラスミドDNA pAL79と,それを持たないプラスミドpUC19を,大腸菌JM107株を用いて調製し,その構造を比較した。電子顕微鏡観察の結果,負に超らせん化し たpAL79 DNAにおいては,複数の分子が集合した構造を示すものが観察されたが,pUC19 DNAにおいては,そのような凝集構造は見られなかった。クロロキンを用いた二次元電気泳動の結果から,pAL79におけるDNA-DNA相互作用は,負 の超らせんの解消にともなって消失(凝集構造の解離)することが示された。その結果,ミニサテライトDNA領域においては,負の超らせん環境下でDNA- DNA相互作用を提供する何らかの高次構造形成が予測された。この構造の詳細を明らかにするために,DNAの塩基特異的化学修飾実験や,より高分解能の電 子顕微鏡観察を進めている。
また,我々はこれまでに,分子内三重鎖構造等のDNA特殊高次構造を,単離したDNA分子を用いて,電子顕微鏡により可視化しているが,これらの特殊高次構造の存在を細胞核内クロマチン構造上で検出することについて,種々の予備的実験を行っている。
【追加情報】(調査月2007年8月、記入月2008年6月)
発表論文
瀬尾芳輝,若松永憲(獨協医科大学 医学部)
鷹股 亮(奈良女子大学 生活環境学部)
荻野孝史(国立精神・神経センター 神経研究所)
森田啓之(岐阜大学 大学院医学研究科)
吉本寛司(京都府立医科大学 大学院医学研究科)
村上政隆(生理学研究所)
我々は,脳機能画像法(fMRI)により,自律神経中枢の神経活動を空間的・経時的に測定し解析する事 を目標として実験を行っている。2005年度は,高張のNaCl溶液を投与(高浸透圧負荷)し,その結果引き起こされる視床・視床下部・延髄における体液 や循環などの自律神経機能中枢の反応を経時的に追った。側脳室内に高張のNaCl溶液を投与すると,SFO,OVLT,LHA領域に急速な反応が見ら れ,3分間遅れてPVNやSONに神経活動の増加に伴う反応が検出された。同条件下に腎交感神経活動を測定すると100秒程度で増加する速い神経活動の増 加と9分程度必要とするゆっくりした神経活動の増加が見られた。この結果から,SFOやOVLT等の脳室周囲器官は,脳脊髄液の浸透圧増加を即座に検知し PVNやSONに伝達する(速い反応)と共に,おそらくPVNやSON自身も脳室から緩やかに伝わってきた高浸透圧を検知する(遅い反応)ものと考えられ る。
さらに細かい神経活動を追うには,低侵襲にMnイオンを脳組織にローディングする必要がある。我々は, 脳室内投与,脳実質内投与,血管内慢性投与,腹腔内投与等の様々なプロトコールを検討した。脳室内投与は,脳室に比較的近い神経核における経時的変化を捉 えるに有利であり,複数回の刺激にも対応した神経活動の検出が可能である。しかし,深部の核の測定,あるいは脳室近くの核と遠い核との同時測定には,なか なかよい負荷条件を見いだせなかった。一方,腹腔内投与では,経時的な情報は追うことができないが,均一にMnイオンを負荷できるので,脳全体の広空間分 解能三次元画像により,反応部位のスクリーニングを行うのに適していることがわかった。我々の実験結果では,一度Mnイオンで造影しても比較的短時間 (24-48時間)で,神経細胞外に排泄されるという結果がえられた。繰り返しの実験が可能となり好都合であるが,同様の方法を用いた研究報告では数日か ら1週間にわたり造影効果が持続したとあり,大きな差がある。おそらく,Mnイオンの投与法の違い,それによる神経毒性の軽減の結果,我々の場合短い造影 時間が得られたものと思われる。
なお,神経ペプチドやホルモンなどの効果については,現在データを収集中である。
【追加情報】(調査月2007年8月、記入月2008年6月)
関係論文
杉谷博士,吉垣純子,松木美和子(日大松戸歯)
橋本貞充(東京歯大)細井和雄(徳島大大院医歯)
林 知也(明治鍼灸大柔道整復)恵良聖一(岐阜大学医分子生理学分野)
村上政隆(統合バイオ生理研)
分泌腺からの最終分泌液に含まれる蛋白質は腺細胞が合成する内因性のものと血液より移行する外因性のも のの混合物である。最近,唾液蛋白の分析が新しい質量分析器により進んだが,内因性と外因性を分離しての研究は未着手である。これには細胞接着を通過する 傍細胞輸送調節の理解が必要であった。本研究は,内因性蛋白の分泌調節に関わる実験系と外因性蛋白分泌をモニターする実験系を作成し,分泌顆粒放出機構と 傍細胞輸送機構による蛋白sortingについて検討した。
1) 我々は以前,顎下腺におけるシグナル伝達として,唾液分泌刺激となるムスカリン性受容体の活性化の下流シグナルとしての一酸化窒素 -cGMP系の存在を明らかにした。今回は,ウサギ顎下腺におけるcGMP分解調節について検討したところ,11種類のcyclic nucleotide phosphodiesterase (PDE) ファミリーのうちPDE1およびPDE2がムスカリン性受容体刺激によるcGMP分解に関与することを明らかにした。
2) 現在のところ,開口放出機能を維持した唾液腺の培養細胞の存在は認められていない。そこで,ラット耳下腺腺房細胞の初代培養条件の検討を行った。 Collagen-Iでコートした培養皿にて分離耳下腺腺房細胞を培養したところ,形態的には変化したものの,48時間目までb- およびムスカリン性受容体刺激による開口放出が認められた。また,初代培養耳下腺腺房細胞においては,新規な分泌顆粒の合成や,遺伝子導入時には新規な開口放出関連タンパク質の発現も認められ,唾液腺における開口放出研究に有用な手段と考えられる。
3) 唾液腺における水チャンネルとしてアクアポリン5 (AQP5) の腺腔側膜での発現は既に認められ得ているが,ラット耳下腺分泌顆粒におけるAQP5の局在を検討したところ,分泌顆粒膜にAQP5が発現していることを 生化学的および形態学的に認めた。抗AQP5抗体で分泌顆粒を処理すると,分泌顆粒の膨張と溶解が認められた。しかし,この抗AQP5抗体の効果は,分泌 顆粒懸濁液のCl-の存在と分泌顆粒膜の透過性に依存していた。これらのことから,AQP5は耳下腺分泌顆粒内浸透圧調節に関与することが示唆された。
4) 唾液腺にはヒトアルブミンの分泌機構が存在しないが,灌流液中タンパク濃度のおよそ0.05 - 0.7%のアルブミンが唾液中に認められた。特徴的な結果として,一酸化窒素(NO)がSH基に結合したS-ニトロソアルブミンが唾液のみに,唾液中アル ブミンのおよそ10-30%の濃度で検出された。NO合成酵素阻害剤であるL-NAMEを灌流液中に加えたところ,唾液分泌量はほとんど変化しない が,S-ニトロソアルブミンはほとんど検出されなかった。これらの結果は,a) 灌流液中のヒトアルブミンがラット顎下腺のタイトジャンクションのような傍細胞経路を通って唾液中に分泌されたこと,b) このタンパク質がその傍細胞経路中でNOを中心とする活性酸素によって修飾を受けたことを示唆している。本実験系は巨大分子が傍細胞経路を通過する過程を 示すものであり,さらにウシアルブミンを用い,唾液分泌速度とタンパク質の唾液中濃度との関係についても検討を開始した。
【追加情報】(調査月2007年8月、記入月2008年6月)
発表論文
関係論文
海老原達彦(産業技術総合研究所)
岡本治正(産業技術総合研究所)
亀山仁彦(産業技術総合研究所)
白幡惠美(山形大学医学部)
早坂 清(山形大学医学部)
中山希世美(日本学術振興会特別研究員)
岡村康司
Cav1.2チャネルに関して,N末端側を欠いたサブユニットを強制発現させたトランスジェニックマウスを作製した。具体的には2種類のマウスを作成した。
1. 他のPSDタンパク同様にbeta-actin promoter 下流にΔCav-GFP をつないだDNA(抑制能はXenopusで確認済み)を導入したトランスジェニックマウス。
2. constitutiveにΔCavを発現すると致死的な恐れがあるため,loxPを用いて普段は"silent"で,Creによって発現が起こるDNA コンストラクトを導入したトランスジェニックマウスを作製した(ΔCav-GFP/Venus)。現在,1のマウスのline化を完了して解析を行ったと ころ細胞内に凝集しているように見えるものの,バックグラウンドと明確に峻別することが出来ず,中断している。ΔCav-Venusについては,別に作製 中のトランスジェニックマウスでVenusが問題なく光る事を確認できたら作製を行う予定である。
以上から,一足飛びにトランスジェニックマウスを作ることが難しい状況であり,ひとまずXenopus oocyteなどの発現系に立ち返り,キメラによる活性化部位同定,その結果に基づくより小さい抑制型のデザインを優先する予定である。
武田 洋幸(東京大学 大学院理学系研究科)
東島 眞一
ゼブラフィッシュの多数のエンハンサートラップラインをスクリーニングすることにより,神経系のきわめ て少数の細胞で発現する転写因子,接着分子,チャネル分子を見付け出し,( i ) 転写因子,接着分子に関しては解剖学的な解析により発生における機能を明らかにし,( ii ) チャネル分子については,電気生理学的な解析により運動系ネットワーク中での生理的な機能を明らかにすることを目標として研究を進めている。今年度の研究 では50系統を超える数のラインについてスクリーニングを行い,中枢神経系の少数の細胞でGFPの発現がみられるいくつかのラインを見いだした。とくに もっとも着目している1つのラインでは,後脳においてはマウスナー細胞特異的に発現がみられ,また,脊髄においては非常に少数(各半体節あたりほぼ1つ) の交叉型介在ニューロンで発現が見られる。この交叉型介在ニューロンは,なにか他とは違う性質をもつニューロンである可能性が高いものと期待される。この ラインの挿入近辺に存在する遺伝子のクローニングを進めるとともに,特異的なGFPの発現がみられる交叉型介在ニューロンについて,電気生理学的解析を 行っていく予定である。
<<最新の採択表に戻る






