変乆偺拞偺乽屆偄擼乿
4寧偵2搙傎偳丄乽恑壔乿偵娭學偡傞彫偝側夛崌偵嶲壛偡傞偙偲偑偁偭偨丅
偦偙偱峫偊偨偙偲傪壓偺昞偵彂偄偰傒傞丅
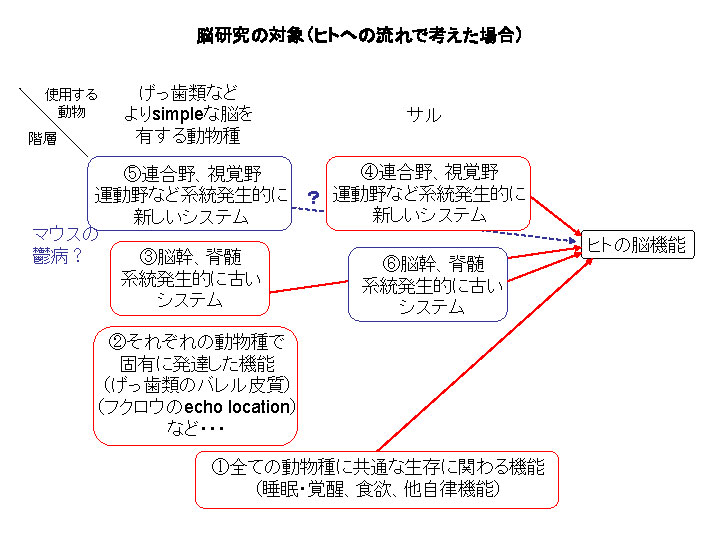
恄宱壢妛偺尋媶偵偍偄偰丄偳偺傛偆側摦暔傪梡偄偰丄壗偺婡擻傪尋媶偡傞偺偐丠偲偄偆偙偲偵偮偄偰丅堦斣壓偵彂偄偰偁傞乽嘆懡偔偺摦暔庬偵嫟捠偡傞惗柦傪堐帩偡傞偨傔偺婎杮揑側婡擻乮悋柊丒妎惲丄怘梸丄懠偺帺棩婡擻乯乿傪尋媶偟偰偄傞尋媶幰偼丄偳偺傛偆側摦暔傪巊偭偰偄偰傕僸僩偵偮側偘傞偙偲丄恑壔丒奒憌惈偺栤戣側偳偱擸傓偙偲偼彮側偄偩傠偆丅傑偨丄乽嘇偦傟偧傟偺庬偵屌桳偵敪揥偟偨婡擻乿傪尋媶偟偰偄傞尋媶幰傕丄媡偵丄僸僩傊偺捈愙偺偮側偑傝傪峫偊側偄強偵傾僀僨儞僥傿僥傿偑偁傞傢偗偩偐傜丄暊傪妵偭偰偟傑偊偽壗傕擸傓偙偲偼側偄丅偦偟偰丄乽嘊斾妑揑僔儞僾儖側擼傪傕偭偨摦暔偱宯摑敪惗妛揑偵屆偄僔僗僥儉乮擼姴丄愐悜丄屆彫擼側偳乯傪尋媶偡傞乿尋媶幰傗丄乽嘋暋嶨側擼傪桳偡傞僒儖側偳偱偄傢備傞擼偺崅師婡擻傪尋媶偡傞乿尋媶幰傕丄偦傟偧傟偺尋媶懳徾偺慖戰偺惍崌惈偵偮偄偰偁傟偙傟尵傢傟傞偙偲傕丄帺恎偑擸傓偙偲傕彮側偄偩傠偆丅堦曽偱丄偦傟埲奜偺晹暘偵偼偦傟偧傟擸傑偟偄栤戣偑偁傞丅傂偲偮偼乽嘍斾妑揑僔儞僾儖側擼傪帩偭偨摦暔乮摿偵儅僂僗側偳乯偱楢崌栰丄帇妎栰丄塣摦栰側偳偺婡擻傪尋媶偡傞乿尋媶偱丄乽偦傕偦傕帩偭偰偄側偄傕偺傪尋媶偱偒傞偺偐丠乿乽儅僂僗偼摑崌幐挷徢丄帺暵徢丄烼昦側偳偺儌僨儖偲偟偰揔愗偐丠乿偲偄偆媍榑偑懡乆偁傝偮偮傕丄幚嵺偵偙偺晹暘偼尰戙偺恄宱壢妛偺椞堟偱偼敋敪揑偵憹壛偟偰偒偰偄傞丅栜榑丄暘巕堚揱妛揑庤朄傪梡偄偰條乆側堚揱巕夵曄摦暔傪嶌傞偙偲偑偱偒傞偺偑嫮傒偩偑丄巹偼偙偪傜偺曽柺偺尋媶偺optimism偼丄慡偰傪斲掕偡傞傢偗偱偼側偄偑彮乆斸敾揑偩丅傗偼傝丄壗傪尋媶懳徾偲偡傞偐偼傛偔峫偊側偔偰偼偄偗側偄丅梫慺偺晹暘偵愗傝暘偗偰夝愅偡傞偙偲偱偼摼傞傕偺偑懡偄偲巚偆偑丄phenoptype偵閤偝傟偰奜憓偑峴偒夁偓偰偟傑偄丄寢壥偲偟偰乽娫堘偭偨乿寢榑偱撍偭憱偭偰偟傑偆偙偲偵側傜側偄偐丄晄埨偱偁傞丅偦傟偵懳偟偰丄幚偼巹偼嵟屻偵巆偝傟偨乽嘐崅搙偵敪払偟偨擼傪帩偮僒儖傪巊偭偰宯摑敪惗妛揑偵屆偄僔僗僥儉傪尋媶偡傞乿晹暘偵懏偟偰偄傞丄偲尵偊傞偐傕偟傟側偄丅偙傟偼埑搢揑偵儅僀僲儕僥傿偩丅幚嵺偵傗偭偰偄傞偺偼愐悜偺恄宱夞楬偩偭偨傝丄屆偄帇妎宯偱偁傞乽旼忬懱奜帇妎宯乿側偳偱偁偭偨傝偡傞丅乽偦傫側偙偲偼儅僂僗傪巊偭偰傗傟偽偄偄偠傖側偄偐丅僒儖傪傗傞側傜丄崅師婡擻両楢崌栰両乿偲埆岥傪尵傢傟偰偟傑偄偦偆偩偑乮幚嵺偵嵟弶偺崰偼傛偔尵傢傟偨乯丄偙傟偵偼巹側傝偺棟桼偑偁傞丅変乆偺擼偼恑壔偺夁掱偱屆偄擼偺晹暘偵怴偟偄僔僗僥儉傪忋忔偣偡傞偙偲偱敪揥偟偰偒偰偄傞丅椺偊偽丄戝擼旂幙偐傜愐悜傊偺搳幩宱楬偼丄壓摍側歁擕椶偩偲愐悜偺夘嵼僯儏乕儘儞傪夘偟偰塣摦僯儏乕儘儞偵愙懕偡傞偑丄崅摍楈挿椶偱偼旂幙愐悜楬偐傜塣摦僯儏乕儘儞傊偺捈愙搳幩宱楬偑弌尰偡傞丅傑偨帇妎宯偵偮偄偰傕丄壓摍側愐捙摦暔傗壓摍側歁擕椶偱偼栐枌偐傜忋媢乮帇奧乯偵搳幩偡傞宱楬偑拞怱偩偑丄崅摍側歁擕椶偱偼栐枌偐傜奜懁旼忬懱傪夘偟偰戝擼旂幙帇妎栰偵帄傞宱楬偑儊僀儞偵側傞丅偙偺傛偆側巹払偺拞悤恄宱宯偺拞偱巆偝傟偨乽屆偄擼丄屆偄夞楬乿偺晹暘偑丄変乆偺乽崅師側婡擻乿乮庤偺岻鉱塣摦丄拲堄丄堄幆etc.乯偺攚屻偱堦懱壗傪偟偰偄傞偺偩傠偆偐丠幚偼偙偺偙偲傪栤戣偵偡傞偲彮側偐傜偸恖偨偪偑彮乆姶忣揑側斀墳傪偝傟傞偙偲傪巹偼宱尡偟偰偄傞丅偙傟偼梞偺搶惣傪栤傢偢偳偆傕偦傟偧傟偺恖偨偪偺尋媶偑棫媟偟偰偄傞崻姴傗丄偦傟偧傟偺悽奅娤丄恖娫娤丄廆嫵娤丄椣棟娤偵懳偟偰憡摉偵僞僢僠乕側栤戣偱偁傞傜偟偄丅偁傞恖偼乽偦偆偄偆屆偄僔僗僥儉偼丄偁偭偰偼搒崌偑埆偄偐傜梷惂偟偰偄傞偺偱偡丅乿偲嫮偔庡挘偡傞丅偙傟偵偼夁嫀100擭梋傝恄宱壢妛偺悽奅傪巟攝偟偰偒偨僪僌儅偺堦偮偱偁傞乽戝擼壔(encephalization)乿偲偄偆峫偊曽偑塭嬁偟偰偄傞偺偐傕偟傟側偄丅偙傟偼壓摍側摦暔偺屆偄擼偺峔憿偑桳偟偰偄傞婡擻偼丄戝擼偺敪払偲偲傕偵戝擼側偳崅師拞悤偵乽堏偭偨乿偲偡傞峫偊曽偱偁傞丅偙偺峫偊曽偵傛傟偽丄戝擼偑攋夡偝傟偨応崌偵偦偺屆偄擼偺婡擻偑尒偊偰偔傞丅尨巒揑斀幩偟偐傝丄戝擼曈墢宯偵撪嵼偡傞忣摦傗杮擻揑峴摦偱偁偭偨傝偡傞丅偙偆偄偆乽屆偄擼乿偺婡擻偑変乆偺摑堦偝傟偨堄幆嬻娫偲偼暿偵丄晛抜偐傜棤偱婡擻偟偰偄傞偐傕偟傟側偄丄側偳偲偄偆偙偲偼懡偔偺恖偵偲偭偰偼梋傝婥帩偪椙偄傕偺偱偼側偄偟丄峫偊偨偔側偄偙偲側偺偐傕偟傟側偄丅
偙偺僪僌儅偺晛曊惈偵偮偄偰偺媍榑偼偝偰偍偒丄乽巹払偺擼偺拞偺屆偄僔僗僥儉乿傪尋媶偡傞棫応偲偟偰丄巹偼傑偢怴偟偄僔僗僥儉偑懝彎偝傟偨応崌偵屆偄僔僗僥儉偑婡擻傪戙彏偡傞丄偲偄偆揰偵偮偄偰偙偺10擭偔傜偄尋媶偟偰偒偨偑丄偙傟偵偮偄偰偼偁傞掱搙偺惉壥傪廂傔傞偙偲偑偱偒偨丅旂幙愐悜楬偐傜塣摦僯儏乕儘儞傊偺捈愙宱楬傪懝彎偡傞偲愐悜夘嵼僯儏乕儘儞宯傪夘偡傞宱楬偑戙彏偡傞偟丄堦師帇妎栰傪懝彎偡傞偲忋媢偑婡擻傪戙彏偡傞丅
偦偙偱師偺僞乕僎僢僩偼丄傗偼傝丄偦傟偱偼偙傟傜乽屆偄夞楬乿偑晛抜変乆偺擼偺拞偱壗傪偟偰偄傞偺偐丠偑栤戣偩丅怴偟偄擼偵傛偭偰梷惂偝傟偰偄傞偺偐丄偦傟偲傕悘帪婡擻偟偰偄傞偺偐丠偙傟偵偮偄偰偼丄椺偊偽Action blindsight偲屇偽傟偰偄傞尰徾偱偼丄TMS偱堦師帇妎栰傪堦帪揑偵梷惂偟偰scotoma傪嶌偭偰丄帇妎揑堄幆傪憆幐偝偣偰傕丄儕乕僠儞僌塣摦拞偵僞乕僎僢僩偺応強傪媫偵偢傜偟偨嵺偵榬偺婳摴傪捛悘偟偰嬋偘傞丄偲偄偆婡擻偼巆傞偲偄偆曬崘傕偁傞乮Christensen MS et al. PNAS, 2008乯丅偙偺傛偆側榬偺婳摴曄峏偑柍堄幆偵婲偒傞偲偄偆偺偼婛偵1980擭戙偐傜Goodale傜偵傛偭偰曬崘偝傟偰偄傞丅偙偺寢壥偼丄柍堄幆偺姶妎乗塣摦宯偲偟偰忋媢側偳偑晛抜偐傜婡擻偟偰偄傞壜擻惈傪帵嵈偟偰偄傞丅偟偐偟丄偙傟傑偱僷儔僟僀儉偼乽怴偟偄擼乿傪梷惂側偄偟偼攋夡偡傞偙偲偵埶偭偰偄傞偙偲偑彮乆栤戣偩丅
巹偼幚尡壢妛幰偩偐傜丄偙偺傛偆側乽屆偄僔僗僥儉乿偺婑梌偵偮偄偰偼傗偼傝媍榑偡傞偩偗偱側偔丄幚尡揑偵徹柧偟側偔偰偼偄偗側偄丅偦偙偱栶偵棫偮偩傠偆媄弍偼変乆偑崱僒儖偵揔梡偟偰偄傞optogenetics偲偐僂傿儖僗2廳姶愼朄偵傛傞摿掕恄宱夞楬偺宱楬慖戰揑丄壜媡揑揱払幷抐朄偩丅偙傟偵傛偭偰変乆偺擼偺拞偺乽屆偄僔僗僥儉乿傪壜媡揑偵幷抐偡傞丒丒丒偙傟偵偮偄偰偼彮偟寢壥偑弌偰偒偰偄傞偺偱岊偆偛婜懸両
乽偛垾嶢2011擭4寧乿丂乽偛垾嶢2011擭3寧乿丂
乽偛垾嶢2011擭2寧乿丂乽偛垾嶢2011擭1寧嘦乿丂乽偛垾嶢2011擭1寧嘥乿丂乽偛垾嶢2010擭12寧嘦乿丂乽偛垾嶢2010擭12寧嘥乿
乽偛垾嶢2010擭11寧嘦乿丂乽偛垾嶢2010擭11寧嘥乿丂乽偛垾嶢2010擭10寧嘦乿丂乽偛垾嶢2010擭10寧嘥乿丂乽偛垾嶢2010擭9寧乿
乽偛垾嶢2010擭8寧嘦乿丂乽偛垾嶢2010擭8寧嘥乿丂乽偛垾嶢2010擭7寧乿丂乽偛垾嶢2010擭6寧乿丂乽偛垾嶢2010擭5寧乿
乽偛垾嶢2010擭4寧乿丂乽偛垾嶢2010擭3寧乿丂乽偛垾嶢2009擭12寧乿丂乽偛垾嶢2009擭11寧乿丂乽偛垾嶢2009擭6寧乿
乽偛垾嶢2009擭3寧乿丂乽偛垾嶢2009擭1寧乿丂乽偛垾嶢2008擭12寧嘦乿丂乽偛垾嶢2008擭12寧嘥乿丂乽偛垾嶢2008擭11寧乿
乽偛垾嶢2008擭9寧乿丂乽偛垾嶢2008擭4寧乿丂乽偛垾嶢2008擭2寧乿丂乽偛垾嶢2007擭12寧乿丂乽偛垾嶢2007擭11寧乿
乽偛垾嶢2007擭10寧乿丂乽偛垾嶢2007擭8寧乿丂乽偛垾嶢2007擭7寧乿丂乽偛垾嶢2007擭6寧乿丂乽偛垾嶢2007擭4寧乿
乽偛垾嶢2007擭3寧乿丂乽偛垾嶢2007擭1寧乿丂乽偛垾嶢2006擭4寧乿丂乽偛垾嶢2006擭1寧乿丂乽偛垾嶢2005擭5寧乿
|